The true power of Carnotaurus sastrei
Dec 20, 2021 1:53:39 GMT 5
Life, theropod, and 2 more like this
Post by Infinity Blade on Dec 20, 2021 1:53:39 GMT 5
Hello. I wanted to make a brief review covering the functional anatomy of the theropod dinosaur Carnotaurus sastrei, with my own conclusions about the animal based on what I know. We currently lack a dedicated profile for this species, and I wanted to focus on some particular aspects of the animal, hence why I opted to make a thread conveying my own viewpoints. Carnotaurus has sometimes been referred to as a "dinosaurian cheetah". I noticed throughout my time on the Internet that this has led to it being characterized as a physically "unimpressive" animal (id est, in terms of physical strength and weaponry), like the modern cheetah. I disagree: several aspects of Carnotaurus' anatomy (and that of abelisaurids in general) point to a rather powerful, impressive, and physically imposing animal.
1.) Hypertrophied caudofemoralis musculature:
Carnotaurus has been found to have had a massive Musculus caudofemoralis muscle in its tail, even relative to other theropod dinosaurs (Persons & Currie, 2011).
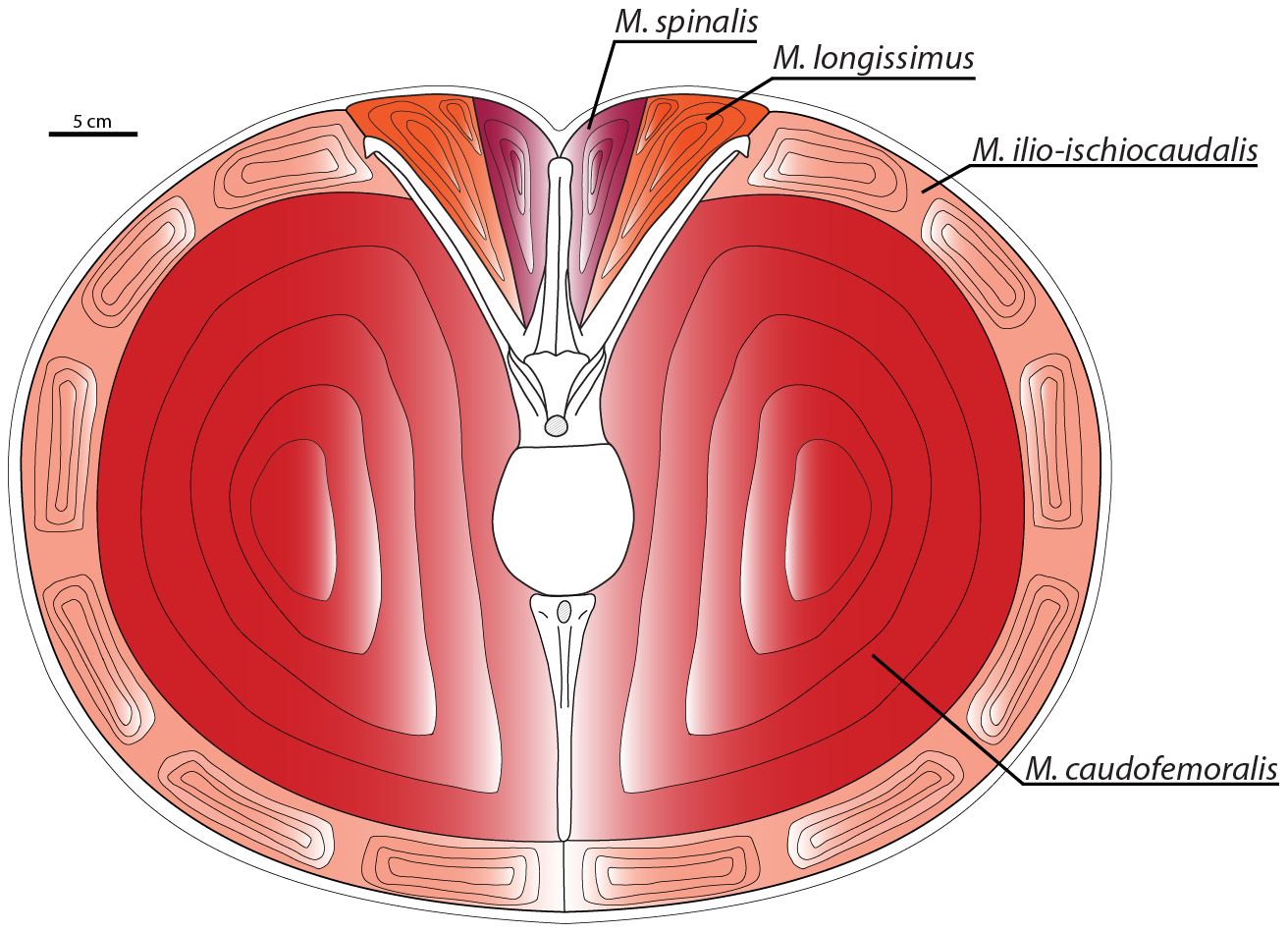
Fig. 5 of Persons & Currie (2011). Cross section of the caudal musculature of Carnotaurus. Notice the extremely enlarged M. caudofemoralis.
The primary interpretation of this feature is that Carnotaurus was potentially cursorial, since the massive M. caudofemoralis granted it great power to propel itself forward while running in a straight line (it should be noted that most of the lower legs are still unknown, so to what extent it was cursorial is therefore also unknown). But by extension (because this muscle would be employed for any forward locomotion), this muscle would also add extra power when the animal was wrestling or pushing forward against another animal (e.g. pushing against a rival with its horned head; more on that below) or attacking “explosively”, giving it a powerful forward thrust.
Conclusion: the hypertrophied M. caudofemoralis muscle would have granted great power to Carnotaurus as it pushed/shoved/wrestled head-on against other animals, and increased its overall physical strength.
2.) Powerful calf muscles:
The cnemial crest is a bony muscle attachment to tibial muscles. The larger and more projected the cnemial crest is, the larger the muscles it can have attached to it are, and the more leverage they have. Abelisauroids in general are typified by a large, hatchet-shaped cnemial crest (Novas, 2009).
Known elements of Carnotaurus stop just below the knee where the cnemial crest is (see this skeletal-> by Jaime A. Headden for known material of Carnotaurus). This fact, coupled with modern skeletal reconstructions, and the typical condition for abelisauroids to have well developed cnemial crests, seem to suggest that Carnotaurus had large, well-developed cnemial crests, and therefore powerful shank musculature.

Skeletal reconstruction of Carnotaurus by randomdinos->
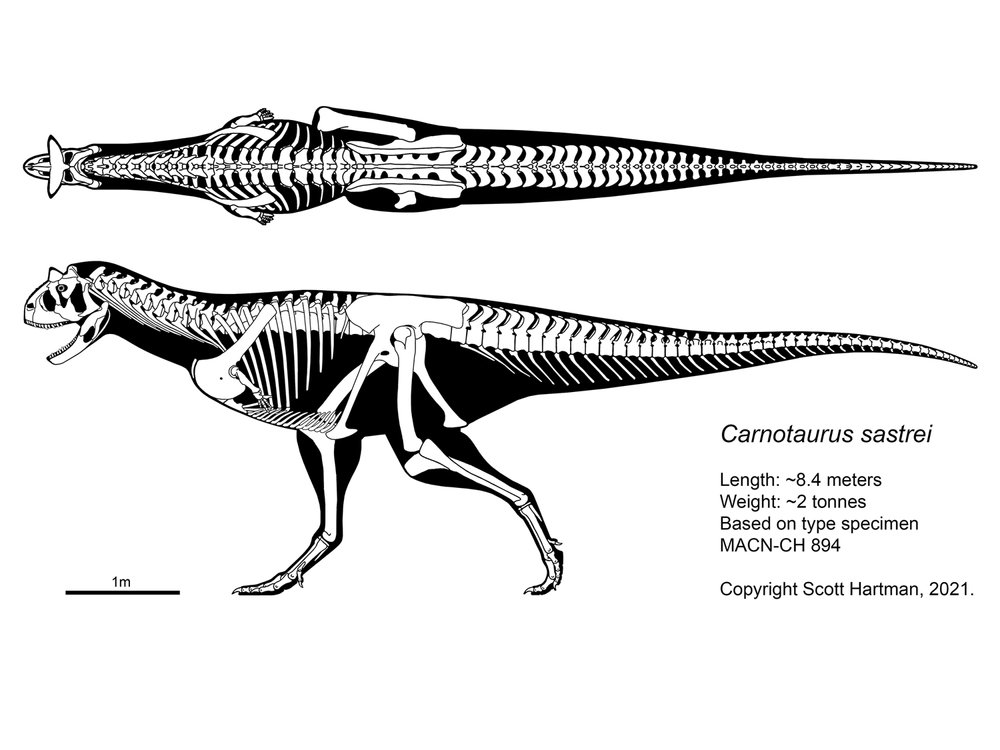
Another skeletal reconstruction of Carnotaurus by Dr. Scott Hartman.
Just like the enormous tail muscles, these powerful lower leg muscles would have increased its overall physical strength, especially as it was pushing and wrestling against other animals by producing strong ground-reaction forces with its legs, or as it made an “explosive” attack.
Conclusion: Carnotaurus seems to have had powerful calf muscles that would also have served to increase its overall physical strength, which would be especially useful in the context of pushing/shoving/wrestling head-on.
3.) Powerful neck muscles:
Reconstructions of Carnotaurus’ neck musculature (based on muscle scars present on the bone, as well as comparison to modern birds and crocodilians) suggest its neck musculature was as proportionately massive as that of a rhinoceros (like the Indian rhinoceros), and twice as massive as that of a hippopotamus (Mazzetta et al., 1998).

Additionally, the pectoral girdle would have augmented the attachment of powerful neck musculature. Some neck muscles would have attached to the scapula and likely contributed to feeding (Burch, 2017), and the enlarged coracoid (shared with other abelisaurids) possibly provided further attachment for enlarged neck musculature (Novas, 2009).
As stated above, this powerful neck musculature would be useful in feeding (Burch, 2017), as well as in handling and dismembering prey (Delcourt, 2018).
Strong neck musculature would also have been useful for withstanding powerful blows with the head. Mazzetta et al. estimate that Carnotaurus’ epaxial musculature was able to withstand impacts (from another rival) of up to 5.7 m/s (Mazzetta et al., 1998).

Novas (2009) also corroborates this.

Conclusion: Carnotaurus had very powerful neck muscles that would have been useful for both handling and dismembering prey, as well as absorbing certain amounts of shock.
4.) Rigid vertebral column:
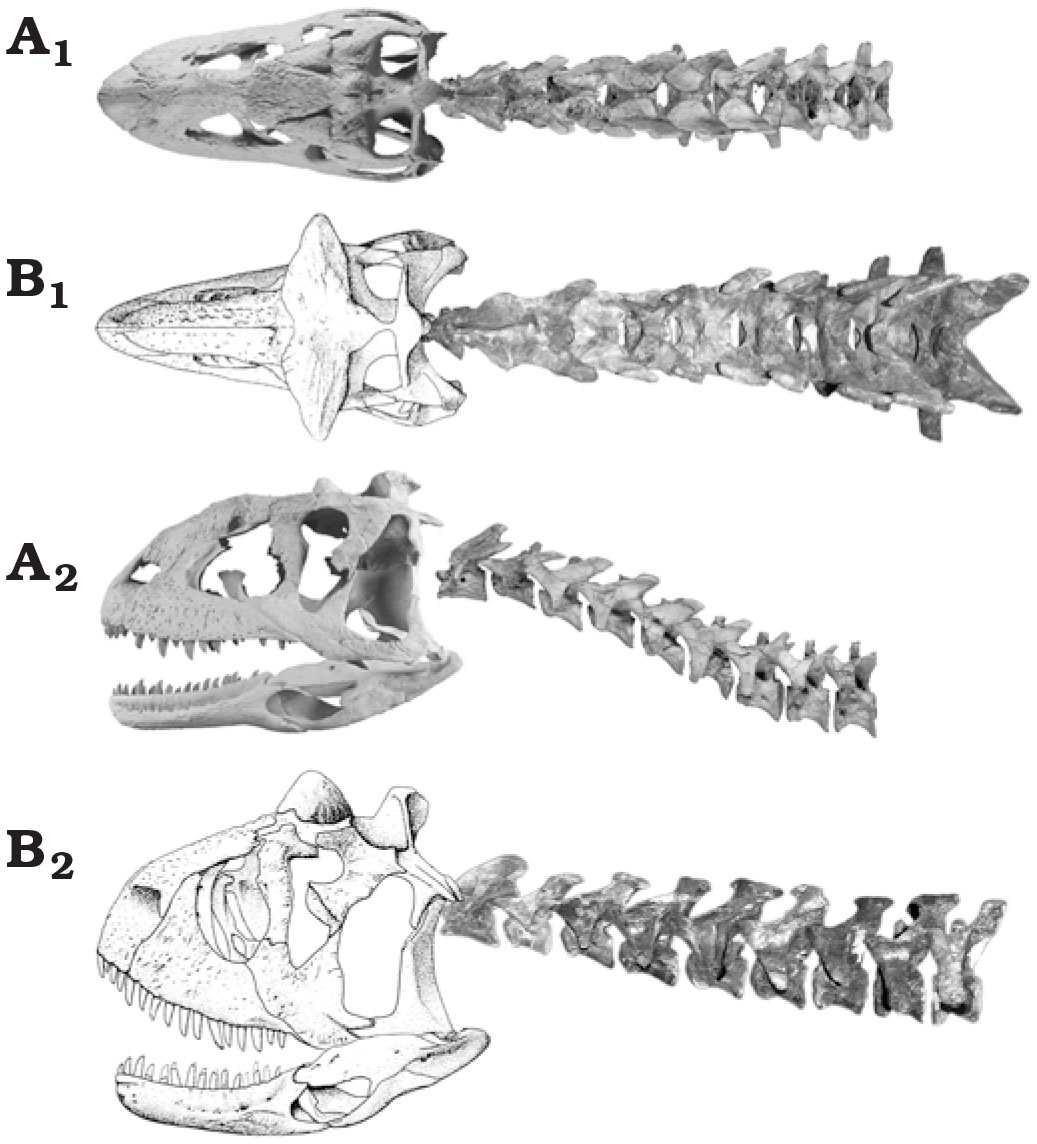
Carnotaurus belongs to a clade of abelisaurids called the Furileusauria (“stiff-backed lizards”). As their name suggests, these abelisaurids are distinguished by their stiff axial column (Filippi et al., 2016).
This would likely function to counter violent shocks (Novas, 2009).

Conclusion: Carnotaurus had a stiffened post-cervical vertebral column that would have been well-braced for shock absorption (to a certain extent).
5.) Powerful bite and slicing teeth:
Previous studies have attempted to calculate the bite force of Carnotaurus, but the most recent biomechanical analysis of theropod mandibles (using finite element analysis) found that the holotype specimen of Carnotaurus (mandible length of 50.3 cm) seems to have had a bite force of 10,735.1 Newtons (=2,413 lbf) (Rowe & Snively, 2021; Table 6). This is much more powerful than a previous figure of ~3.3 kN (Mazzetta et al., 2009). This study appears to have measured actual bite reaction forces at teeth (particularly two mesial teeth), so this should be actual bite force.
The highly sculpted pattern of the cranial bones suggest hypermineralization of the skull’s soft tissues. This, coupled with the thick and strong skull bones, would have helped relieve tension when biting (Cerroni et al., 2020).
It should be noted that Carnotaurus did not have cranial kinesis, or at least not to the extent proposed by Mazzetta et al. (1998). Nor was its mandible “extremely weak” as it was described long ago (Cerroni et al., 2020).
As is typical for predatory dinosaurs, Carnotaurus’ teeth were still blade-like, ziphodont instruments that were suitable for cutting flesh. In fact, the teeth of abelisaurids such as Carnotaurus are even more compressed labiolingually than those of carcharodontosaurids (Candeiro et al., 2012).
Conclusion: Carnotaurus appears to have had a powerful bite, with adaptations to help diminish stress during the bite. This, coupled with the highly labiolingually compressed ziphodont teeth, would have given it a damaging bite.
6.) Horns:
Carnotaurus is perhaps best known for the horns on top of its head (this is, in fact, what gave the creature its name, which translates roughly to “meat-eating bull”). The skull was sufficiently well built to withstand low-impact headbutts and pushing and shoving loads (though not high-impact ramming at maximum speed) (Mazzetta et al., 2009). Giraffe-like strikes to the necks and flanks of rivals are also possible (Delcourt, 2018). Overall, use of the skull and horns in some form of agonistic behavior seems to be their most likely function (Cerroni et al., 2020). Whether such horns were used against prey or intraspecific competition (or perhaps even both) is uncertain.
Conclusion: Carnotaurus had a skull and horns that could be used for pushing/shoving, as well as low-impact headbutts and/or giraffe-like blows to flanks.
Overall conclusion:
In contrast to its image as a “dinosaurian cheetah” (which was never meant to be taken literally, and is seemingly based solely on its speed), Carnotaurus actually possessed a suite of characteristics that made it a physically powerful and dangerous animal. In summary:
1.) The femur-retracting tail and leg muscles were very powerful, offering it significant strength against other animals.
2.) The neck muscles were also very powerful, giving it great ability to handle/dismember prey and absorb shock when using the head/horns as a weapon. The atrophied forelimbs likely even gave way for increased neck musculature attaching to the pectoral girdle.
3.) Most of the vertebral column seems to have been stiffened, strengthening its back. This would be useful when physically wrestling and for absorbing shock.
4.) As it turns out, the bite seems to have been powerful and, coupled with slicing teeth, substantially damaging.
5.) The skull and horns seem to have been useful for combat, particularly for striking and pushing (though not for ramming).
With its rather rigid back and tail vertebrae (Novas, 2009; Persons & Currie, 2011; Filippi et al., 2016), Carnotaurus was not a theropod especially suited for agility (although, the powerful leg and tail muscles would probably still give it sheer brute force when making turns). Instead, it seems to have been very well suited for a frontal engagement with its musculature, strengthened back, and weaponry. Coupled with a body mass of ~1,500 to ~1,600 kg (Mazzetta et al., 1998; Benson et al., 2014), Carnotaurus would have been a physically powerful, well-armed, gigantic predator.
1.) Hypertrophied caudofemoralis musculature:
Carnotaurus has been found to have had a massive Musculus caudofemoralis muscle in its tail, even relative to other theropod dinosaurs (Persons & Currie, 2011).
Digital muscle models indicate that, relative to its overall body size, Carnotaurus sastrei had a substantially larger M. caudofemoralis than any other theropod yet studied. In most non-avian theropods, as in many extant sauropsids, the M. caudofemoralis served as the primary femoral retractor muscle during the locomotive power stroke. This large investment in the M. caudofemoralis suggests that Carnotaurus sastrei had the potential for great cursorial abilities, particularly short-burst sprinting.
Fig. 5 of Persons & Currie (2011). Cross section of the caudal musculature of Carnotaurus. Notice the extremely enlarged M. caudofemoralis.
The primary interpretation of this feature is that Carnotaurus was potentially cursorial, since the massive M. caudofemoralis granted it great power to propel itself forward while running in a straight line (it should be noted that most of the lower legs are still unknown, so to what extent it was cursorial is therefore also unknown). But by extension (because this muscle would be employed for any forward locomotion), this muscle would also add extra power when the animal was wrestling or pushing forward against another animal (e.g. pushing against a rival with its horned head; more on that below) or attacking “explosively”, giving it a powerful forward thrust.
Conclusion: the hypertrophied M. caudofemoralis muscle would have granted great power to Carnotaurus as it pushed/shoved/wrestled head-on against other animals, and increased its overall physical strength.
2.) Powerful calf muscles:
The cnemial crest is a bony muscle attachment to tibial muscles. The larger and more projected the cnemial crest is, the larger the muscles it can have attached to it are, and the more leverage they have. Abelisauroids in general are typified by a large, hatchet-shaped cnemial crest (Novas, 2009).
The abelisauroid tibia is distinguished by a large cranioproximally projected hatchet-shaped cnemial crest, as exemplified by Quilmesaurus…
Known elements of Carnotaurus stop just below the knee where the cnemial crest is (see this skeletal-> by Jaime A. Headden for known material of Carnotaurus). This fact, coupled with modern skeletal reconstructions, and the typical condition for abelisauroids to have well developed cnemial crests, seem to suggest that Carnotaurus had large, well-developed cnemial crests, and therefore powerful shank musculature.
Skeletal reconstruction of Carnotaurus by randomdinos->
Another skeletal reconstruction of Carnotaurus by Dr. Scott Hartman.
Just like the enormous tail muscles, these powerful lower leg muscles would have increased its overall physical strength, especially as it was pushing and wrestling against other animals by producing strong ground-reaction forces with its legs, or as it made an “explosive” attack.
Conclusion: Carnotaurus seems to have had powerful calf muscles that would also have served to increase its overall physical strength, which would be especially useful in the context of pushing/shoving/wrestling head-on.
3.) Powerful neck muscles:
Reconstructions of Carnotaurus’ neck musculature (based on muscle scars present on the bone, as well as comparison to modern birds and crocodilians) suggest its neck musculature was as proportionately massive as that of a rhinoceros (like the Indian rhinoceros), and twice as massive as that of a hippopotamus (Mazzetta et al., 1998).
Additionally, the pectoral girdle would have augmented the attachment of powerful neck musculature. Some neck muscles would have attached to the scapula and likely contributed to feeding (Burch, 2017), and the enlarged coracoid (shared with other abelisaurids) possibly provided further attachment for enlarged neck musculature (Novas, 2009).
The retention of large scapulocoraoids in nonavian theropod taxa with reduced limbs may be due to a close developmental association between the scapular blade and the axial skeleton (Kuijper et al. 2005; Valasek et al. 2011; Dececchi & Larsson, 2013), but some of the muscles attaching to the scapulocoracoid possibly had important roles in other activities. Those muscles attaching to the neck (e.g. Levator scapulae) could have played a part in feeding, as may be the case in extant crocodylians (Meers, 2003), and those muscles attaching to the ribs (e.g. Serratus muscles) could have had a role in respiration, as they appear to have in extant birds and crocodylians (Codd et al. 2005; Munns et al. 2012).
The abelisauroid shoulder girdle is characterized by a strap-like scapular blade and an unusually wide elliptical-shaped coracoid, a condition verified in Carnotaurus, Majungasaurus, Aucasaurus, Indosuchus, and the basal abelisauroid Deltadromeus (Fig. 6.11). The enlarged coracoid seems incongruent with the pronounced reduction in size of the entire forelimbs. The wide coracoidal surface may instead reflect an enlargement of the area for attaching muscles of the neck rather than those muscles that inserted on the atrophied forearms.
As stated above, this powerful neck musculature would be useful in feeding (Burch, 2017), as well as in handling and dismembering prey (Delcourt, 2018).
According to the analyses of Therrien et al., carnotaurines (e.g. Majungasaurus and Carnotaurus) might have been ambush predators attacking large prey. Additionally, Sampson and Witmer have suggested that Majungasaurus, and possibly other carnotaurines, were “adapted for a mode of predation that entailed relatively few, penetrating bites accompanied by powerful neck retraction, as well as bite-and-hold behaviour”. This predatory behaviour is consistent with results on skull biomechanics as well as neck analyses.
Strong neck musculature would also have been useful for withstanding powerful blows with the head. Mazzetta et al. estimate that Carnotaurus’ epaxial musculature was able to withstand impacts (from another rival) of up to 5.7 m/s (Mazzetta et al., 1998).
Novas (2009) also corroborates this.
Conclusion: Carnotaurus had very powerful neck muscles that would have been useful for both handling and dismembering prey, as well as absorbing certain amounts of shock.
4.) Rigid vertebral column:
Carnotaurus belongs to a clade of abelisaurids called the Furileusauria (“stiff-backed lizards”). As their name suggests, these abelisaurids are distinguished by their stiff axial column (Filippi et al., 2016).
Among other anatomical peculiarities, this clade is characterized by axial stiffness, evidenced in the highly developed of structures as epipophyseal processes in the cervicals, hyposphene-hypantrum structures in the dorsals, fused sacral complex, and distal tip of caudal transverse processes.
This would likely function to counter violent shocks (Novas, 2009).
Conclusion: Carnotaurus had a stiffened post-cervical vertebral column that would have been well-braced for shock absorption (to a certain extent).
5.) Powerful bite and slicing teeth:
Previous studies have attempted to calculate the bite force of Carnotaurus, but the most recent biomechanical analysis of theropod mandibles (using finite element analysis) found that the holotype specimen of Carnotaurus (mandible length of 50.3 cm) seems to have had a bite force of 10,735.1 Newtons (=2,413 lbf) (Rowe & Snively, 2021; Table 6). This is much more powerful than a previous figure of ~3.3 kN (Mazzetta et al., 2009). This study appears to have measured actual bite reaction forces at teeth (particularly two mesial teeth), so this should be actual bite force.
This analysis assessed the effects of ligaments, which accommodate more strain than bone, on bone stresses and tooth reaction forces.
To apply forces, nodes were selected in the areas where forces were being tested and divided by the total respective estimated muscle force by the total number of nodes selected. Constraints were assigned to restrict free body motion, at the hinges of the jaw and two mesial teeth for both constraints to obtain bite reaction forces.
The highly sculpted pattern of the cranial bones suggest hypermineralization of the skull’s soft tissues. This, coupled with the thick and strong skull bones, would have helped relieve tension when biting (Cerroni et al., 2020).
Carnotaurus, as well as most abelisaurids (e.g. Abelisaurus; Skorpiovenator; Arcovenator; Majungasaurus) shows postorbital and lacrimal bones with strongly developed dermal projections which almost encloses the orbit. Besides these dermal outgrowths, the mineralisation of the suborbital ligament would have delimited the ventral orbital sector (Chure 1998; Sampson and Witmer 2007). These broad and rough circumorbital bones stands in contrast with the gracile and mostly smooth bones of the basal ceratosaurs Ceratosaurus, Eoabelisaurus and noasaurids (e.g. Masiakasaurus; Carrano et al. 2011). As previously indicated by Sampson and Witmer (2007), abelisaurids maybe had a propensity to mineralise cranial systems; Carnotaurus, as well as Majungasaurus, have an ossified interorbital septum and ethmoidal complex (Paulina-Carabajal 2011a; Paulina-Carabajal 2011b). Further, the calcification of the hyoid bones (ceratobranchials and basihyal) seems to have been influenced by this hypermineralization; these elements usually remain cartilaginous in extant archosaurs, and probably, most dinosaurs. Although the hypermineralization of the skull soft tissues is an abelisaurid feature, it was convergently acquired by carcharodontosaurids and tyrannosaurids (Novas 1997; Holtz 2004), suggesting a propensity on the hypermineralization in many mid- and large-sized theropods. The pattern of highly sculptured cranial bones in concert with the thick and strong skulls, would have diminished tensions during the bite (Henderson 2002; Rayfield 2004); and as proposed for the case of circumorbital bones, it would have protected the eye against eventual blows resulting from intraspecific competition (Hieronymus 2009; Novas 2009).
It should be noted that Carnotaurus did not have cranial kinesis, or at least not to the extent proposed by Mazzetta et al. (1998). Nor was its mandible “extremely weak” as it was described long ago (Cerroni et al., 2020).
In sum, the strong mineralization of most skull roof and circumorbital bones, coupled with coossification of several bone joints, suggest that Carnotaurus lacks cranial kinesis, or at least, it was not as kinetic as was proposed previously by Mazzetta et al. (1998). This supports the results of Sampson and Witmer (2007) for these cranial bones, while a plausible kinesis is restricted to the intramandibular joint.
Although the articulation between dentary and postdentary bones was early described as ‘extremely weak’ (Bonaparte et al. 1990), it should be instead better described as a looser articulation (in terms of ossification) that resulted in a more mobile joint.
As is typical for predatory dinosaurs, Carnotaurus’ teeth were still blade-like, ziphodont instruments that were suitable for cutting flesh. In fact, the teeth of abelisaurids such as Carnotaurus are even more compressed labiolingually than those of carcharodontosaurids (Candeiro et al., 2012).
Cross-sectional shape has already been used to study theropod teeth (Bakker et al., 1988; Currie et al., 1990; Farlow et al., 1991; Sankey et al., 2002), and it is useful for identifying teeth from Brazil. The cross-sections of teeth in the jaws of Aucasaurus garridoi (MCF-PVPH-236), Carnotaurus sastrei (MACN 894), Noasaurus leali (PVL-4061), Indosuchus raptorius (AMNH 1960), and an unidentified Carnotaurinae (UNPSJB-PV-247) are similar to the Peirópolis teeth. Morphotypes one and two have convex lingual and labial faces of the crown, but are compressed along the carina margins. This type of cross-section is different from that of a carcharodontosaurid (Stromer, 1931; DG/UFRJ S/N; RTMP 94.43.1, 94.43.2, 94.43.3, 94.43.4, Giganotosaurus MCPV-CH-1). Abelisaurid (Aucasaurus garridoi, Carnotaurus sastrei and Indosuchus raptorius) teeth are more compressed labiolingually than those of Carcharodontosaurids (Fig. 3). Dentary teeth of Carnotaurus sastrei and Indosuchus raptorius (AMNH 1960) are labiolingually compressed and slightly recurved on the posterior carina whereas in Noasaurus leali and Masiakasaurus knopfleri (Carrano et al., 2002) they are curved more strongly along the posterior carina. Morphotypes one and two have similar characteristics to those in the jaws of Carnotaurus sastrei and Indosuchus raptorius – both have slightly convex lingual and labial faces and compressed edges.
Conclusion: Carnotaurus appears to have had a powerful bite, with adaptations to help diminish stress during the bite. This, coupled with the highly labiolingually compressed ziphodont teeth, would have given it a damaging bite.
6.) Horns:
Carnotaurus is perhaps best known for the horns on top of its head (this is, in fact, what gave the creature its name, which translates roughly to “meat-eating bull”). The skull was sufficiently well built to withstand low-impact headbutts and pushing and shoving loads (though not high-impact ramming at maximum speed) (Mazzetta et al., 2009). Giraffe-like strikes to the necks and flanks of rivals are also possible (Delcourt, 2018). Overall, use of the skull and horns in some form of agonistic behavior seems to be their most likely function (Cerroni et al., 2020). Whether such horns were used against prey or intraspecific competition (or perhaps even both) is uncertain.
Nonetheless, the horns of Carnotaurus are flat on their posterodorsal surfaces (Bonaparte et al., 1990), providing a broad contact surface for the distribution of the impact force during frontal clashes with conspecifics. Moreover, the possibility that the horns may have had a corneous covering (Novas, 1989; Bonaparte et al., 1990), which would have provided them with further cushion to absorb shocks, was not taken into account. Although our constrained model may lead to an underestimation of the safety factor calculated for this activity, the uncertainties regarding the dynamic effect in headbutting combats discouraged us from using more favorable assumptions. Our mechanical analyses support the use of headbutting, provided that they consisted of low-motion, or even quasi-static, contests. Moreover, the strong fusion between frontals and parietals (Bonaparte et al., 1990) should effectively dissipate stresses on the cranium of Carnotaurus, providing increased protection against brain damage during low-motion head-butting contests. If combat did occur, the laterally divergent frontal horns probably engaged such that the contest was settled in most cases by shoving. Such confrontational encounter would have been reminiscent to those seen in bovids as Bos and Bison, but probably did not involve wrestling given the unfavorable orientations of Carnotaurus horns.
Going through the new information of soft tissues of abelisaurids shown here (above), it is possible to infer that this clade might have had some intraspecific headbutting matches behaviour at least in carnotaurine taxa (as suggested for Carnotaurus and Majungasaurus). The presence of cornified cover on the skull, that was inferred for Carnotaurus and Majungasaurus, has been related to headbutting behaviour in extant taxa (e.g. Ovibos moschatus, Syncerus caffer and Buceros vigil) as well as extinct (e.g. Pachyrhinosaurus, Achelousaurus horneri and Stegoceras validum). Nevertheless, differing from those that engage in violent headbutting and have deep cancellous bone (which carnotaurine lack), the carnotaurine might have used the head in low-motion headbutting and shoving matches at low speeds (as marine iguana Amblyrhynchus cristatus) or engaged giraffe-like strikes to each other’s neck and flanks. The giraffe-like strikes have been proposed for Majungasaurus due to the presence of tall, rugose nasals, struts within sinuses and a unicorn-like projection of the frontals, although stresses. Also, the mechanical analyses of Carnotaurus skull performed by Mazzetta et al. support the low-motion headbutting in this taxa. Furthermore, the presence of well-developed occipital region (e.g. nuchal crest) associated with large epipophysis and neural spines in the cervical vertebra increasing the neck musculature strongly suggest that the cervicocephalic complex (head and neck) withstood high stress. Indeed, the well-developed epipophyses indicate a good leverage for intervertebral dorsiflexion by the muscle tranversospinalis cervicis and the origin of a strong muscle complexus, a head dorsoflexior. As similar features on neck and skull are spread throughout the carnotaurine abelisaurs, all the taxa belonging to this clade may have had similar behaviour in territoriality or mating matches for instance. It is worth noting that cranio-facial biting was reported for non-avian theropods. This behaviour could have had several possible reasons, including territoriality, courtship/mating, play, predation/cannibalism, intrapack dominance and subadult dispersal. In the case of carnotaurine, the headbutting and/or giraffe-like strikes could also have been added to the behavioural repertoire for any reasons above.
Many years of discussion and hypothesis were focused on the alleged function of this remarkable trait. Firstly Novas (1989), and posteriorly Mazzetta et al. (1998), proposed that horns would have been used as weapons again other individuals through striking blows with the head, possibly due to intraspecific competition. Also, bony ornaments in theropods were hypothesised as displays for courtship and mating, and as intraspecific recognition through sexual dimorphism (Molnar 2005). However, Hieronymus (2009) proposed that the bony ornaments present in Majungasaurus and abelisaurids functioned to absorb blows in agonistic behaviours (as in extant mammals), due to intraspecific competition. Carnotaurus frontal horns are like those of the bovids, in having a well-developed horn core (Novas 2009). Furthermore, the CT scans information shows that the horns are predominantly solid (see below discussion of pneumatization). The highly developed horns, a thickened skull roof, robust circumorbital bones and a robust neck where the epaxial musculature probably were well developed and capable of absorb shocks (Novas 2009; Méndez 2014), leads to the hypothesis that Carnotaurus would have the potential to use the horns for some kind of agonistic behaviour, or at least, diminish any potential blow. Although, if this behaviour was used again either intraspecific competition or preys, is uncertain. The hypothesis concerning the sexual dimorphism cannot be corroborated because of the solely specimen nature of the holotype.
Conclusion: Carnotaurus had a skull and horns that could be used for pushing/shoving, as well as low-impact headbutts and/or giraffe-like blows to flanks.
Overall conclusion:
In contrast to its image as a “dinosaurian cheetah” (which was never meant to be taken literally, and is seemingly based solely on its speed), Carnotaurus actually possessed a suite of characteristics that made it a physically powerful and dangerous animal. In summary:
1.) The femur-retracting tail and leg muscles were very powerful, offering it significant strength against other animals.
2.) The neck muscles were also very powerful, giving it great ability to handle/dismember prey and absorb shock when using the head/horns as a weapon. The atrophied forelimbs likely even gave way for increased neck musculature attaching to the pectoral girdle.
3.) Most of the vertebral column seems to have been stiffened, strengthening its back. This would be useful when physically wrestling and for absorbing shock.
4.) As it turns out, the bite seems to have been powerful and, coupled with slicing teeth, substantially damaging.
5.) The skull and horns seem to have been useful for combat, particularly for striking and pushing (though not for ramming).
With its rather rigid back and tail vertebrae (Novas, 2009; Persons & Currie, 2011; Filippi et al., 2016), Carnotaurus was not a theropod especially suited for agility (although, the powerful leg and tail muscles would probably still give it sheer brute force when making turns). Instead, it seems to have been very well suited for a frontal engagement with its musculature, strengthened back, and weaponry. Coupled with a body mass of ~1,500 to ~1,600 kg (Mazzetta et al., 1998; Benson et al., 2014), Carnotaurus would have been a physically powerful, well-armed, gigantic predator.