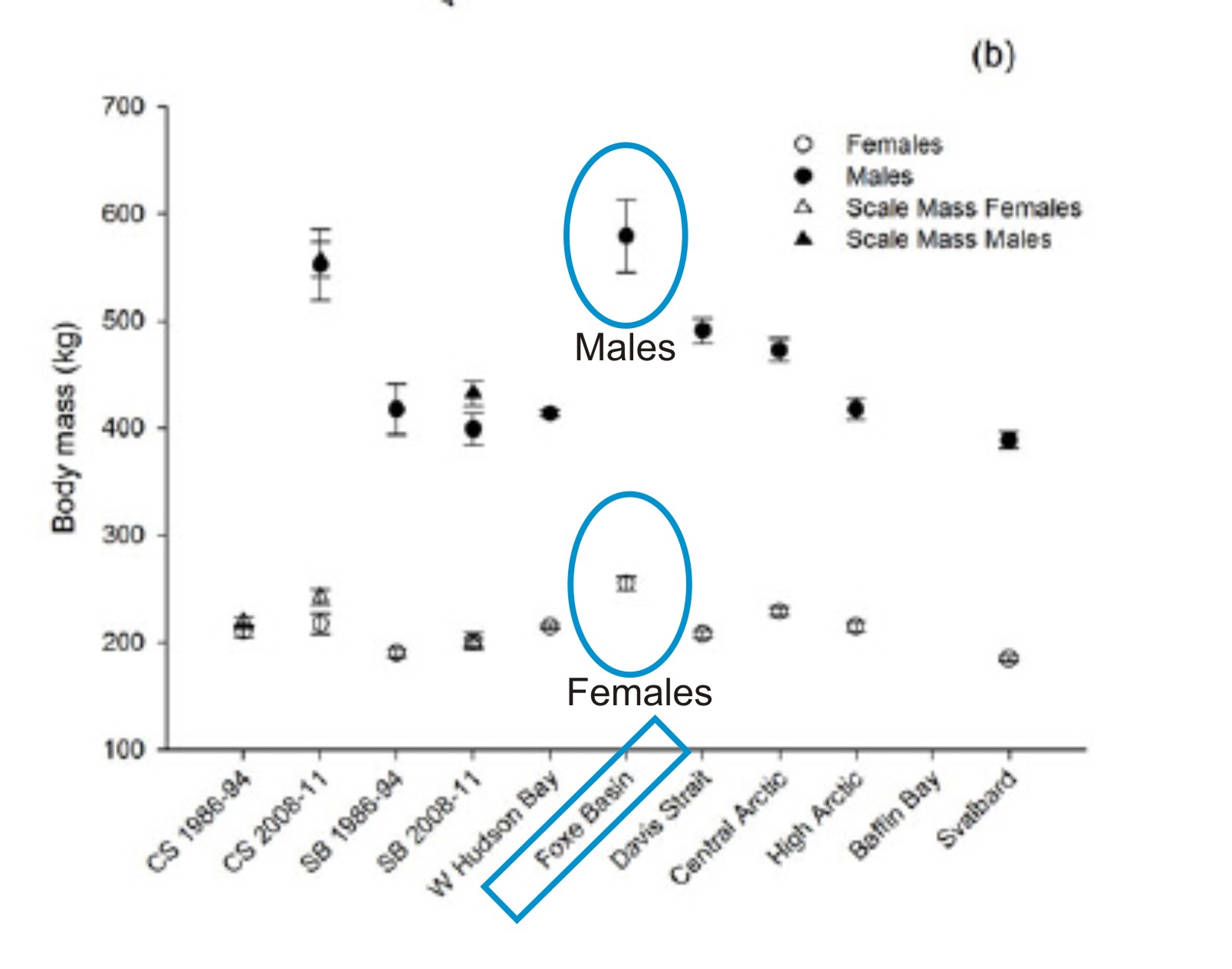
Whether the largest theropod taxa were really all the same size is the point of contention here, so presupposing that they are the same size to argue that there couldn’t be a size difference is still circular.
Isn’t there? I was under the impression that it is pretty much consensus that the siberian tiger is the largest cat, that the kodiak bear is the largest fully terrestrial bear etc.
This extends to other categories too.
That there are often multiple animals reaching broadly similar sizes, or even that in data-deficient situations it is often unclear or harshly debated which is the largest, doesn’t mean there isn’t usually one that can be identified as the largest even among a sample of very large taxa if there are sufficient data.
African and Asian elephants are clearly the largest terrestrial animals in the extant world, but when it comes down to it African Elephants are still clearly larger than Asian elephants, although if this were a point of contention due to statistical uncertainty, someone might argue that they both represent the largest terrestrial animals and so should be expected to be similar in size due to this.
Blue whales are still clearly larger than fin- or right whales. This becomes even more clear once we look throughout earths history. The largest known terrestrial mammals or the largest known terrestrial dinosaurs from different epochs were not all exactly the same size. The largest known marine animals or marine predators weren’t either. In the Devonian, the largest marine predator was a 1 ton placoderm, in the Cretaceous the largest marine predator was a mosasaur likely on the order of 10 tons in body mass, in the Neogene the largest marine predators were a species of giant lamniform shark and a species of giant physeteroid whale that each could probably exceed 50 tons in body mass. There is no reason at all to expect there is some intrinsic or extrinsic limitation that means we have to strongly expect that they were actually all the same size.
The largest theropods throughout the Mesozoic weren’t all the same size either. For all we know, the largest theropods before the late Jurassic weren’t even in the same ballpark, let alone exactly the same size, as
Giganotosaurus or
T. rex. I don’t mean like "they might have been one or two tons smaller than the 6t + that were repeatedly reached by multiple giant theropods from the late Jurassic and Cretaceous", more like "they might have been one or two tons in body mass at most". Expecting that there were multiple lineages of theropods that could have potentially reached similar sizes is one thing. Expecting that every single species among those must have reached exactly the same size and there cannot possibly be one that’s identifiably the largest, or even just identifiably larger than a particular other taxon, is another.
While with some (especially where there’s only one known specimen) it may be impossible to say with confidence which is larger, there is no reasonable expectation that they were all exactly the same, and neither is such an expectation reflected in papers, articles, popular-scientific rankings, documentaries, or on boards like this one. Usually when the known specimens of one taxon imply a larger size than the known specimens of another, that is considered sufficient to expect that it is, more likely than not, larger. That is a standard that most people agree with when it comes to other animals, yet somehow the standard for demonstrating something is larger than
Tyrannosaurus is inexplicably much higher.
To make the claim that both taxon A and taxon B represent the exact upper bound to how large an animal of their respective ecology could get, and that it is impossible for there to be a difference between their sizes, would require a special type of evidence that we do not have.
this statement seems to contradict itself. Either one species is a bit bigger than the next-largest or it isn’t, it can’t be both at the same time.
We don’t actually know if they can. The point is that neither Mallon and Hone, nor I, see any reason to expect that they can’t. Gut feelings that something represents the limit for terrestrial predators aren’t evidence. And based on this framework, it’s frankly not reasonable to expect that while size ranges of individual giant theropod species varied by multiple tons, a taxon of giant theropod cannot possibly be 0.5-1 t heavier than another on average. There is no evidence at all that would preclude this.
What do you consider "fairly common"? As far as the known
T. rex sample is concerned, these are clearly unusually large, which doesn’t mean an absolute rarity when considering the entire population of thousands or millions. Specimens probably would have reached that size with some regularity, but the vast (>90%) majority of the adult specimens you would find at any given time would have been smaller.
Yes, and specimens of other theropods that were dramatically bigger also probably existed, and there’s just as little telling if they will ever be found. But the point being, in taxa that likely had intraspecific size ranges (for adult specimens) between 4 or 5 and 15 tons, there is nothing outlandish about the possibility that one taxon may be a bit bigger than the other on average.
A 2.5 m tall human suffering from a developmental condition is not a reasonable equivalent to one species of giant theropod from an entirely different lineage perhaps being 10% heavier than another species of giant theropod.
This is a caveat to any such "agility index" that treats the body as a rigid beam, but I think I’ve already discussed that. Of course in real life, giant theropods could and would have also flexed parts of their axial skeleton to improve turning ability.
I know, that’s another thing I have already discussed here, and that’s one reason why, as I explained, the agility index gives an incomplete picture, because it only includes the area of the ilium and not other relevant musculature that is situated in the tail.
That’s the point. There isn’t, and nobody I know has ever argued it either, yet based on the same metric that Snively et al. calculated for Tyrannosaurids and Allosauroids (limb muscle origin size/rotational inertia), you would clearly have to expect that they can.
The point is not that you could directly compare the agility index for a quadruped to that of a biped. You can’t, as I already explained, but for our purposes here there is no need to either. However you can compare them between two different quadrupeds with similar bauplan, just as you can (that’s what Snively et al. did after all) compare them among different bipeds with similar bauplan.
As you’ll note I actually did modify the equation to apply to quadrupeds, by also including the area of the scapula and not just the ilium. Otherwise it could be validly argued that perhaps a difference would be due to different relative forelimb and hindlimb muscle sizes (just as it can be argued that at least part of the difference between tyrannosaurids and allosauroids might be due to different relative tail and iliac muscle sizes).
I didn’t say that it was, but this happens with such frequency when it comes to
T. rex that I’m strongly inclined to think that in this context, it is.
The majority of arguments frequently brought forth in favor of
T. rex, are arguments that, when they apply to other animals, are rarely if ever made or taken seriously. Brain size, metabolism (at least if it’s not a downright massive endotherm vs ectotherm scenario), binocular vision and top speed are all things that are frequently portrayed as notable, perhaps even major advantages in this scenario, but very rarely even discussed in any other analogous scenarios.
I don’t even recall having ever seen a discussion of relative brain sizes of canids or felids, for example, or of the relative widths of their fields of stereoscopic vision. I didn’t check, but they are unlikely to be exactly the same between all mammalian predators. Differences that in other animals get summarily dismissed as irrelevant, or not even looked at in the first place, are suddenly interpreted as major advantages as soon as they apply to
T. rex.
The rotational inertia/agility index is just one example of that. And while a specific feature can certainly be argued to be more relevant in some contexts than in others (which doesn’t necessarily mean that I agree that it is in this case), if this reasoning is almost always applied to all features it does indicate a double standard, or in this case, a systematic bias that has become so pervasive it must be considered a cultural phenomenon.
Sure, but the context difference here is that one is a predator-prey situation and the other is a confrontation between two predators which are both adapted to hunt and kill large animals.
Nobody would even think twice about whether a wildebeest would be a match for hyena at the same size, while a jaguar and a hyaena, or two giant theropods for that matter, will naturally be a lot more closely matched at equal sizes. You can say for the leopard vs wolf analogy (once more, never intended as an all-round analogy for this fight, merely as an example to point out the culturally pervasive and internalized double standards when it comes to
T. rex) what you will, but at least it’s a comparison between two similar-sized hypercarnivorous macropredators, not between a predator and it’s natural prey.
That context difference (predator vs prey as opposed to predator vs predator) does not exist between the analogue I brought up and this scenario. That doesn’t mean there is no difference in contexts or that you cannot argue it is relevant, but in this case I don’t share your opinion that it is, at least not for the validity of the point I was making with the analogy in question (that things are routinely and systematically interpreted as advantages, often major ones, for
Tyrannosaurus that nobody would think of much of they applied to some other animal).
I have multiple issues with this.
First, and most importantly, that’s not all the research there is, and once more it paints an incomplete picture, because the only giant allosauroid it considers is
Meraxes, whereas Allosauroids exhibited fairly disparate growth strategies.
While it’s true that in the specific example you cite,
T. rex was shown to have had a higher peak growth rates (due to a different growth strategy, acceleration vs hypermorphosis) than
Meraxes, Tyrannosaurids in general likely did not have higher growth or metabolic rates than Carcharodontosaurids.


From D’emic et al. 2023. You’ll note that the highest growth rate in this sample (which includes
T. rex) was in
Mapusaurus, a carcharodontosaurid and the closest known relative of
Giganotosaurus.
So for what it’s worth, there is no reason at all to suspect that
Giganotosaurus had lower growth rates than
Tyrannosaurus, although indeed closely related carcharodontosaurids appear to have differed markedly in their growth strategies. Note that differences in growth strategy among related taxa are not at all unheard of (not just among theropods, as you can see above). E.g. stem-physeteroids apparently grew and matured much more quickly back in the Pliocene (Gilbert et al. 2018) than extant toothed whales with similar ecology do (this was interpreted to be the result of predation pressure necessitating a faster growth strategy, though the explanation doesn’t really matter much here). It’s at best very doubtful if these differences imply comparable differences in adult metabolism (I.e. when growth has already slowed or stopped anyway), let alone mobility (were Pliocene physeteroids much more metabolically active and athletic than extant orcas just because they had shorter lifespans and grew more quickly? Unlikely. The same goes for tyrannosaurids vs carcharodontosaurids, except when viewed to a lens of already expecting that tyrannosaurids must have been "more advanced" and "more warm-blooded", which admittedly is what many people do).
Secondly, growth rates are a poor proxy for the metabolic metric that is actually relevant for mobility, which is
maximum metabolic rate, not basal metabolic rate. A more useful proxy to look at would be femoral blood flow index (see Seymour et al. 2012) which correlates more strongly with MMR, and thus gives a better idea of the metabolic metric that is relevant for athletic capacity, but I don’t think that has ever been quantified for giant theropods. For example, elevated maximum metabolic rate is the reason extant varanids have elevated stamina and athletic capabilities despite having low, typically reptilian basal metabolic rates (and something similar probably applied to many basal Synapsids, see Knaus et al. 2021).
Thirdly, this is not just possible, but highly likely. For one because at the time of adulthood there was no more such difference in growth rates (if anything, the Carcharodontosaurid in the graph you posted was growing more quickly than the Tyrannosaurid at time of death, although both were very clearly growing negligibly by that point), but also since basal metabolic rate generally scales at a power of body mass less than 1 (see da Silva et al. 2006), so the larger animals become, the slower their metabolism. Since it was my impression we were comparing adult specimens, not fast-growing subadults, that’s highly relevant. A fast growth rate (whether that’s what we see in a carcharodontosaur or a tyrannosaur, that’s still fast when you compare it to a crocodile) implies an endothermic metabolism because otherwise such high growth rates would never be possible, but that doesn’t mean every tachymetabolic animal necessarily grew at the same speed (obviously, otherwise fully grown specimens would have to turn into ectotherms in order to stop growing). A fully grown 8 ton
T. rex almost certainly had a lower basal metabolic rate than a 3 ton
T. rex that was still in the middle of its growth spurt, but it also certainly still had a much higher BMR than an ectothermic crocodilian of similar body mass and (since the tyrannosaur would be effectively done growing by that point) growth rate.
This also shows you one more caveat;
T. rex almost certainly also had a lower metabolic rate than
Gorgosaurus or
Daspletosaurus, because it was a much larger animal. However, it simultaneously had much higher peak growth rates, because it (unlike some other theropods that had different growth strategies) achieved its gigantism by accelleration, not hypermorphosis. Comparing
Meraxes to Sue in this regard is comparing apples to oranges comparison, because Sue was most likely close to twice the body mass of the
Meraxes type (which has a femur circumference of 452 mm according to Canale et al., compared to Sue’s 580 mm), so of course even with the same growth strategy, it would have had higher peak growth rates to reach its larger body size. In fact on this account,
Meraxes might well have had the higher metabolic rate despite its slower growth, simply on account of being a smaller animal.
For what it’s worth, Barrick and Showers 1999 (ya know, the nineties and early 2000s, those brief few years when
Giganotosaurus was almost as hyped, and frequently the model organism of random case studies, as
T. rex is now…), the one study I am aware of that actually directly analyzed
Giganotosaurus fossil material with regard to thermophysiology (in this case using isotope proxies for homeothermy) and compared it to
T. rex, found no evidence it was any less endothermic than the latter. On the contrary:
They suggest intermediate metabolic rates between modern reptiles and modern mammals for both of these theropods (also more evidence for metabolic rates decreasing in adult individuals as opposed to young ones);
There are a few issues I have with this study that are off topic here (and that I think I’ve already addressed elsewhere years ago), but it did analyze both
T. rex and
Giganotosaurus and found nothing at all to suspect the former had higher metabolic rates, so that’s that (and still probably the most direct information on this specific question that currently exists in the literature).
We sure do, especially since there’s no basis at all to the claim that Carcharodontosaurs in general had lower metabolisms or even slower growth than Tyrannosaurids.
Be that as it may, I think all of it goes to show that it’s one more example of what I keep noticing; nobody would look at two different mammalian predators, or even at a shark and a whale (where there most certainly is a metabolic difference, but not one that many people would say really matters), and think of comparing their growth rates in order to establish an advantage for one and the other.
But when it comes to
Tyrannosaurus, suddenly this is exactly what people do. It just seems like grasping at straw to coax out any imaginable advantages for
T. rex over other theropods, valid or otherwise, things that nobody would ever even consider relevant for other animals. We saw this with brain size, with binocular vision, with turning ability, with growth rates. The double standard here is not arguing that perhaps one feature is more relevant in one scenario than in another, it is systematically considering features as amazing advantages in this specific scenario that are never even discussed in any other even half-way comparable scenarios.
I have said before that the tyrannosaur bias doesn’t spare published research, although obviously that doesn’t mean it invalidates research findings because this bias is expressed very differently, but where it is really rampant is in science journalism and (even more) in the online paleo enthusiast community. Every single "we’ve identified a cool feature of
Tyrannosaurus/tyrannosaurids" paper (of which there are many, but not because they fundamentally have more cool features, but because the ones they do receive way more attention and research interest than they would in another animal) gets totally blown out of proportion and overinterpreted through the "
T. rex was superior"-lens (you can tell, for example from earlier, unqualified and unsourced comments on this very thread that simply stated the belief that
T. rex "seems more warm-blooded" and therefore superior without a need to bring evidence of such claims). Where in research, the bias is mostly expressed in the kinds of studies that get done vs the ones that don’t, the amateur community tends to jump to the conclusion that differences in research interest automatically reflect a difference in interesting biological features.
This is why we get a study on tyrannosaurid rotational inertia and ilium area, but not a study about lateral ranges of hindlimb motion or about lateral spine flexibility, things that would be more likely to run in favor of non-tyrannosaurid theropods. And the picture that paints in the non-professional community is that there is some proportionality between the number of interesting or unique morphological adaptations that get studied and the number that exist, but this is not the case.
This is a misleading size comparison, since that (like pretty much all comparisons) is showing a very large
T. rex skull. The difference would not look as stark as that between a cat and similar-sized canid if one made an attempt to show a more average-sized
T. rex, but it would look quite a bit more noticeable than it does here. Anyway, that there is a vast difference between the skull size of a canid and felid, but not between an allosauroid and tyrannosaurid, was exactly my point. This is a point that goes strongly in favour of the dog (which is also the more agile one, at least based on its agility index), but does not go in favour of the Tyrannosaurid.
Exactly.
Well, I personally don’t agree.
That’s your prerogative, but the source didn’t really say, note or analyze anything that I am not equally qualified to tell you. Cau simply noted that the dorsally angled femoral heads in Carcharodontosaurs would have allowed for more stability by enabling its legs to adopt a wider stance (something plainly obvious from the pertinent osteological descriptions). On a more general note, this (wider limb ranges of motion outside the parasagittal plane) fits the bill of what you would expect when you compare the limbs of more vs less cursorial animals; the former generally tend to have more restricted ranges of motion. This has neither been quantified nor extensively discussed, on Cau’s blog or elsewhere, and it likely never will. Although it it was an advantage of the Tyrannosaur, I’m sure there’d be a paper on it by now and a few hundred online enthusiasts mentioning it every time scenarios like this one get discussed.
I would argue the reverse, especially with a size advantage for the carcharodontosaur.
??
Bite force would obviously have been far greater towards the back of the jaws than the front, all computational simulations show this (the difference is by nearly a factor of two). The most important biting positions would have likely been in the mid-maxillary region, where the largest teeth are, not towards the front (where there are small, incisiform teeth with a d-shaped cross-sections, interpreted to have primarily served for the purposes of defleshing carcasses).
Clearly, but equally clearly theropods were unable to do deathrolls, and postcranial movements would have been important for both allosauroids and tyrannosaurids, not uniquely the latter.
Not if the forces applied are equal.
It doesn’t. Try pricking yourself on the tip of your geological hammer, then try the same with the tip of your pocket knife. The latter will penetrate your skin with vastly greater ease. The hammer of course can apply more force, but that is not a property or advantage of its tip.
Picks are better at penetrating (mostly through armor) because they have far more force behind them than a thrust with a sword does, which is mostly a result of their top-heavy balance. If you stab something with sword and then stab the same thing with the spike of a pick and measure the force it takes to penetrate each time, the sword will penetrate more easily (because it is sharper and thinner), at least given that it’s a relatively pointy, stab-centric sword and not something designed with an extremely blunt tip (like certain falchions).
This advantage of the pick mainly comes in when armor penetration is desired, and is pretty irrelevant against soft targets. Picky weren’t used much against unarmoured opponents not just because of overpenetration (you can overpenetrate far further with a sword, since it has a much longer blade), but because they don’t do as much damage against a soft target, and because they are more cumbersome to wield due to the same qualities that make them good at delivering forceful blows to armor. Hence why swords are usually preferred to axes/maces/picks in unarmoured combat; they tend to be better at injuring soft targets (at least as good at penetrating, vastly better as slashing), and are much more nimble due to how they are balanced.
In this scenario, both are clearly unarmoured.
Giganotosaurus’ jaws and teeth are a better equivalent to a sword (or an even better one to a macuahuitl or a samoan war club), those of
T. rex are a better equivalent to a warhammer. Yes, the latter will penetrate better against an armoured target because they are inherently less fragile and have more force behind them, but against soft tissues (the kind that all vertebrates have around their skeleton), the former will be more damaging.
That depends on how well someone sharpens their sword, the majority used in actual warfare likely weren’t better cutters than shark or varanid teeth are. Of course you can technically sharpen (good quality) steel to be sharper than any tooth, but in practice most swords aren’t sharpened nearly that much. I recently got into blacksmithing, and believe me, getting and maintaining a razor edge on something the length of a sword isn’t easy. Many professional militaries in the modern period didn’t bother with it at all, and kept their sabres in metal scabbards that would dull the edge when pulling it out.
No.
Based on Smith et al. 2005, the teeth of
Carcharodontosaurus have a slightly higher average crown length (mean of 78 mm) than those of
Tyrannosaurus (70 mm). If we exclusively compare maxillary teeth,
T. rex’ teeth are marginally longer on average, but by less than 1 mm (79 vs 78 mm):

There are very few Giganotosaurus teeth preserved with the holotype, none from the upper jaw, and none have reported crown measurements, so, I can’t add them here, but the dentary teeth (Coria and Salgado 1995, fig. 1) appear to be longer than any known
T. rex tooth crown, even though the specimen is smaller than many of the largest
T. rex individuals that are routinely included in studies (and even though the dentary teeth in both tyrannosaurids and carnosaurs appear to usually be somewhat shorter than the maxillary ones). I’m admittedly not 100% sure that there is no root slippage there, that’s why we would need measurements taken on the actual fossil to be confident about how the tooth lengths compare.
Saying that tooth crown lengths of
T. rex were generally greater, than those of giant carcharodontosaurids, let alone enough for it to matter, seems to be unfounded, probably a result of people being used to seeing casts of Stan everywhere (thanks BHI for-profit fossil collecting!), and of tooth lengths for isolated specimens being often reported with the root included (in which case of course
T. rex teeth are of course much longer, but that’s not functional length that would be relevant to penetration).
Yes (force, not necessarily pressure), but they will also require much more force to penetrate (i.e. achieve the same pressure), so listing that as an advantage of the teeth (being better for penetration) is highly misleading. The teeth are indeed worse for penetration, requiring more force, which is only counterbalanced by more force being available.
Higher bite force probably being the single most frequently listed, discussed and agreed upon advantage of
Tyrannosaurus against basically any other theropod it’s ever been compared to, I don’t think there is a need to discuss the point that T. rex, obviously, had the stronger bite. But there is the tendency to try to find ways to sneak it into lists of advantages multiple times, which effectively turns one advantage counterbalanced by a disadvantage (=stronger bite but blunter teeth) into two advantages (=stronger bite and teeth better for penetration).
For what it’s worth, I’ve handled both, and clouded leopard canines actually appear to be sharper than
T. rex teeth (although carina sharpness on mammalian teeth is a lot less uniform than in reptiles, since it depends on fine intraspecific variations of tooth occlusion and wear, so this is probably not the same for all individuals).
I don’t think spiked weapons are any more prone to overpenetrating than swords are (a warhammer or bec du corbin cannot overpenetrate too much, its spike is simply not long enough to penetrate much further than what would be intended and also needed to reliably dispatch an opponent), although I could see how a sword (due to having sharp edges) may be easier to remove from a wound than a blunt pick or spike, because it can effectively "cut its way out" of a wound, something a blunt weapon obviously cannot (that is an advantage cutting teeth share as well, an animal with a cutting bite will have an easier time letting go when it wants to).
Because there are no extant, multi-ton, terrestrial carnivores.
The causative factor here seems unclear. Is that because they have horns or tusks, or simply because of their size? You seem to favor the explanation that it is because they have horns or tusks…but for an elephant or rhino, it is equally advantageous to gore its opponents flank (and potentially inflict lethal penetrating trauma) as it is for a predator that relies on biting to get past the snout and bite a more vulnerable area. I think they are mostly less inclined to maneuver while fighting because locomotory strategies that are open to smaller animals aren’t as easily available to them, making it more effective to just stand their ground, and use their weapons and brute strength to try and gain an advantageous position.
Indeed.
This would still be difficult, if not near-impossible, for a large theropod to pull off against another large theropod, even if we assume there is a major difference in turning ability between the two, because doing so would require not just grappling ability but also to be able to move a lot more than the other animal would have to in order to reposition itself to fend off such an attempt.
The question is what position maneuvering could get either animal into that would actually represent an advantage for it.
For example, assume two large theropods somehow ended up maneuvering alongside each other, the way the rhinos do in this video. Which is absolutely something I could see happening in confrontations between giant theropods as it would be much more quickly accomplished (and thus happen more easily, whether intentionally or accidentally) than one animal turning its body by 90 degrees one way and then back to meet the other with body axes perpendicularly.
In this scenario it doesn’t really matter which one has the better turning ability and maybe initiated that maneuver, it would still represent an advantageous position for the carnosaur, which would most likely be able to inflict critical damage more quickly and easily (large gape + slicing teeth against the unprotected flanks or thighs of the opponent) than the reverse. The ability to capitalize on an advantageous position gained from maneuvering would appear to largely be contingent on the suitability of the weaponery and to an extent on reach and striking speed that would optimize the range of targets that could be attacked in such a situation, and the speed with which they could be attacked. Also potentially on strength (mainly a function of body mass) if it devolves into any kind of shoving or grappling match.
The most surefire way for an animal to provoke a fall is sharp maneuvering or collisions during fast locomotion. Except for household accidents maybe.
Isn’t locking jaws basically the primary argument for why people think
Tyrannosaurus’ jaws to be superior? I am well aware that face-biting is primarily a non-lethal, intraspecific combat strategy, and likely wouldn’t be the winning move in a fight to the death (that would be tearing out the throat), this is the reason why I think an advantage in this regard is somewhat overrated. But it is the key point in which
Tyrannosaurus is argued to hold an advantage, because it has the stronger bite force, which would make it better suited to biting a hard, bony target like the snout. So if we agree that is probably not the most sensible target in a fight to the death, that should make our opinions align more on this.
Sue would have an agility index approx 40% greater than the
Giganotosaurus holotype. More at equal weights, on the other hand probably less if a proxy for the size of the the caudofemoralis was included in the index rather than just the size of the ilium (which is apomorphically expanded on the lineage leading to extant birds, where the caudofemoralis is completely reduced).
That sounds like a lot, but that’s where it is important to put it into perspective by noting that a wolf also has an agility index approximately 40% greater than a similar-sized leopard, something that in practice nobody even notices.
It’s important because the agility index is an index, not a physical quantity (like the absolute rotational acceleration these animals were capable of) that can be usefully interpreted on its own. It tells us no more and no less than the component that body shape and the area of the considered limb muscle origins have in regard to the animals’ agility, but it doesn’t tell us the absolute turning speeds of these animals, nor does it tell us about any other factors influencing it in a living animal (of which there are many, as I’ve listed before; caudofemoral musculature not considered by Snively et al., limb muscle moment arms, muscle tissue, lateral flexibility/ranges of motion all could have major impacts that are unaccounted for).
But where exactly is the evidence to say that
Tyrannosaurus was capable of faster linear acceleration than similar-sized Carnosaurs?
Running down, not outmaneuvering.
So? Elephants are quadrupeds with a short, compact body. They are obviously able to make far tighter turns than 11-12 m long theropods (even those with fancy enhanced turning ability, like tyrannosaurids) would, and especially, they are much more stable while doing it and therefore have a substantially lower risk of falling or other injury than a biped would.
Yet I also don’t generally see elephants circling and trying to outflank each other during fights. You might argue that the two elephants in question would have very similar levels of turning ability, and that is of course correct, but two giant theropods fighting each other also probably wouldn’t have read Snively et al. 2019 in preparation for their fight, and wouldn’t know that one of them might be able to turn more quickly than the other. In fact, based on usual hunting experience against compact quadrupeds like
Triceratops or
Ankylosaurus, one should expect that a
T. rex would not be likely to instinctively assume it can out-turn an opponent and hence try to capitalize on that, even if it theoretically could.
But if it is not going to ram into it at high speeds, that makes it unlikely it will be able to ever even reach its top speed in such a scenario. Accelerating and decelerating to and from this speed would take quite a bit of time for such a large animal. The speeds the animals would practically reach during such a confrontation thus would have much less to do with theoretical top speeds than with acceleration, and being more cursorial doesn’t automatically entail faster acceleration at all.
Approaches at high speed, especially those involving fast maneuvering, do not seem cautious to me with regard to the risk of falling. It has previously been asserted that maintaining body equilibrium is a key constraint to the athletic capacity of a giant theropod (Blanco and Mazzetta 2001). Coupled with the time it would take a theropod of this size to even reach its top speed (which is unlikely to differ by a truly major amount anyway), that makes me doubt it would realistically ever be a factor in a confrontation between two giant theropods, unless it devolved into a chase where both could take their time to accelerate to the highest possible speed they could muster without anticipating having to immediately decelerate again.
Ultimately I think in no small part the predominant opinion on this matchup comes down to frequent misrepresentations (always comparing the most complete and well-known specimens for both, ignoring that for
T. rex, that means using the largest specimens while for
Giganotosaurus it is using the smallest).
When most people think of this scenario, they envision something like the larger red silhouette below being compared to the blue silhouette, and naturally they conclude that
T. rex is much chunkier, larger, more powerful, has larger teeth, a larger brain etc., but that’s because they are comparing very much non-representative individuals, a bit like comparing a huge human bodybuilder smallish gorilla and deciding that hmm, maybe humans are actually as large as or larger larger than gorillas. A quick google search for "T. rex Giganotosaurus" shows that this is the case, you almost exclusively get bombarded with size comparisons between MUCPv-Ch1 and Sue, very rarely some other (above average-sized)
T. rex specimen, not a single
T. rex specimen that was below average in size. I wonder what the mainstream opinion of this interspecific comparison would be if in every size comparison you see online, Sue was replaced with Black Beauty.
A more accurate representation would be the smaller red silhouette, which corresponds to a roughly average-sized
Tyrannosaurus, and even then the
Giganotosaurus should probably be made slightly larger too (and made somewhat more robust, because of the incorrectly restored shoulder girdle in this old reconstruction), because the depicted specimen is still only the holotype. Looked at under these conditions, many of the
T. rex’ advantages suddenly shrink into insignificance, while much about the
Giganotosaurus (e.g. its skull) suddenly looks a lot more impressive once they are being compared fairly.

–––
Barrick, R.E. and Showers, W.J. 1999. Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus. Palaeontologia Electronica 2 (2): 22pp.
Blanco, R.E. and Mazzetta, G.V. 2001. A new approach to evaluate the cursorial ability of the giant theropod Giganotosaurus carolinii. Acta Palaeontologica Polonica 46 (2).
Calvo, J.O. and Coria, R. 1998. New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the largest theropod ever found. Gaia 15: 117–122.
Canale, J.I., Novas, F.E. and Pol, D. 2015. Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) from the Lower Cretaceous of Patagonia, Argentina. Historical Biology 27 (1): 1–32.
Canale, J.I., Apesteguía, S., Gallina, P.A., Mitchell, J., Smith, N.D., Cullen, T.M., Shinya, A., Haluza, A., Gianechini, F.A. and Makovicky, P.J. 2022. New giant carnivorous dinosaur reveals convergent evolutionary trends in theropod arm reduction. Current Biology 32 (14): 3195-3202.e5.
da Silva, J.K.L., Garcia, G.J. and Barbosa, L.A. 2006. Allometric scaling laws of metabolism. Physics of Life Reviews 3 (4): 229–261.
Gilbert, K.N., Ivany, L.C. and Uhen, M.D. 2018. Living fast and dying young: life history and ecology of a Neogene sperm whale. Journal of Vertebrate Paleontology 38 (2): e1439038.
Knaus, P.L., van Heteren, A.H., Lungmus, J.K. and Sander, P.M. 2021. High Blood Flow Into the Femur Indicates Elevated Aerobic Capacity in Synapsids Since the Synapsida-Sauropsida Split. Frontiers in Ecology and Evolution 9.
Smith, J.B., Vann, D.R. and Dodson, P. 2005. Dental morphology and variation in theropod dinosaurs: Implications for the taxonomic identification of isolated teeth. The Anatomical Record 285 (2): 699–736.
Seymour, R.S., Smith, S.L., White, C.R., Henderson, D.M. and Schwarz-Wings, D. 2011. Blood flow to long bones indicates activity metabolism in mammals, reptiles and dinosaurs. Proceedings of the Royal Society B: Biological Sciences 279 (1728): 451–456.