|
|
Post by creature386 on Jun 30, 2015 19:38:07 GMT 5
Even though the second paper is not peer reviewed, it pretty much sums up my opinion. Kicking would only be possible if one is on the ground and forelimb fighting also must have been very hard. However, given that Peck and Jane are two species out of many with facial scars, it is possible that they were the ones where the exception was recorded.
|
|
Deathadder
Junior Member
 aspiring paleontologist. theropod enthusiast.
aspiring paleontologist. theropod enthusiast.
Posts: 240 
|
Post by Deathadder on Jun 30, 2015 20:20:18 GMT 5
I think that while hunting they would place there foot on the sides of the prey to help push it down
|
|
|
|
Post by Infinity Blade on Jun 30, 2015 20:33:58 GMT 5
Even though the second paper is not peer reviewed, it pretty much sums up my opinion. Kicking would only be possible if one is on the ground and forelimb fighting also must have been very hard. However, given that Peck and Jane are two species out of many with facial scars, it is possible that they were the ones where the exception was recorded. It could simply have been that the individual with the pathologies had its head in a lowered down position (e.g. feeding from a carcass or drinking water), only to get attacked. There is, of course, also the possibility that the kicking tyrannosaur was lying on its back and lashed out with its legs like a cat (although, how/why the tyrannosaur got on its back is up for questioning). And, as you mentioned, the kickee itself could have been on the ground (although, I don't know how/why it would get there in the first place; wrestled down? Sleeping beforehand?). And even though the rebuttal argued that tooth marks can vary with shape, a pic in the book I gave a link to shows that a Tyrannosaurus pedal claw fits perfectly within the puncture holes. We can't ignore this: it would be one helluva coincidence if a tooth managed to create a whole that a claw just so happens to snugly fit inside. I decided to co-address the paper (I say 'co-address' because a guy already addressed the main points in the comments, which the author did not effectively counter at all IMO). The aforementioned guy said: [To add to the point about the hindlimbs, the attacking tyrannosaur did not necessarily even have to be standing. A kick could have been delivered by a tyrannosaur lying on the ground, lashing out with its hindlimbs like a cat. In such a case, the issue of stability completely vanishes.] And now, the author's reply: This doesn't at all explain the non-parallel state of the punctures; if a tyrannosaurid bit multiple times and shook its head/neck like a dog, it would create parallel puncture holes given the fixed state of the teeth; it's not like it had movable jaw parts. Multiple bites in the same area would also produce even more bite marks that would overlap, but that's not what we see at all. Also, since when does bone deform in the way proposed here? If a tyrannosaurid shook its head while biting onto a conspecific's face, with sufficient force the teeth would either slip out of the punctures they created (and possibly create a furrow in the bone in the direction they slid off in), break off, or maybe even bite off a piece of bone. Bone is a hard material. It's not going to simply deform by having a hole expand as a puncturing object moves around in it, as it would with say, a thin piece of plastic. Maybe because the particular situation called for it (say, if the tyrannosaur's face was close to its opponent's torso or something)? Maybe to use a less lethal/damaging means of combat? To, you know, minimize risk of serious injury to themselves? It's not unprecedented for animals to use secondary, less deadly weapons in intraspecific combat. Look at male sperm whales; they can ram each other with those massive heads of theirs powered by their >40 tonne bodies swimming at maximum speed, but they also bite each other's faces and create raking wounds with their teeth. (Forelimb usage is not my main concern here, though, I'm more focused on the pedal claws) With all due respect, this is the part of his reply that by far falls the shortest. First, Manning et al. (2006) merely said that dromaeosaurid claws didn't function as cutting implements (and it should be noted that an accipitrid-esque stabbing-to-death function is likely for dromaeosaurid digit ll pedal claws), which is not really what was proposed for the claws of tyrannosaurids (although, it should be noted that modern terrestrial theropods have been known to cause lacerations with their claws/kicks for whatever it's worth). The second sentence seems to be a non sequitur; what does the fact that Rothschild mainly used mammal claws as examples have anything to do with how tyrannosaurids couldn't have done so? Lastly, in regards to claw sharpness. Even if tyrannosaurid foot claws were blunter than those of other theropods (which I question looking at pictures of tyrannosaurid claws and those of other theropods online), that doesn't mean tyrannosaur hind foot claws are blunt altogether or ineffective weapons. Look at pictures online of tyrannosaurid foot claws; those that lack what seems to be taphonomic wear still have clearly defined points at their tips. It's like saying that because the horns of American plains bison are blunted that they're not very useful for offense; the truth is that an American bison can create deep gashes on each other’s necks with their horns. Hell, even claws that are truly blunt can cause damage. Brown bears wear their claws out from locomotion and digging, but they can still cut through each other's skin and expose the flesh underneath. Ratite claws get worn from touching the ground, but they can still lacerate or even disembowel humans with kicks from their clawed feet (although, ostrich claws do still have some pointed tip to them, just not the wear facet itself). Camelids and even equids can do the same with their hooves (which are homologous to claws). |
|
|
|
Post by Infinity Blade on Aug 26, 2015 7:24:33 GMT 5
|
|
|
|
Post by theropod on Aug 26, 2015 16:17:49 GMT 5
I don’t really think so, the claw cross-sections they figured don’t look any sharper than those of dromaeosaurs (as I mentioned before, those also have a sort of edge), which are themselves usually thought of as too blunt for cutting (e.g. Carpenter 1998, Manning et al. 2006). Of course there are variables like the shape of the keratin sheath and the size and strength of the involved animals and tissues, but going by shape alone they don’t seem more suitable for it.
–––References:
Carpenter, Kenneth (1998): Evidence of predatory Behaviour by carnivorous Dinosaurs. Gaia, 15 pp. 135-144.
Manning, Phillip L.; Payne, David; Pennicott, John; Barrett, Paul M.; Ennos, Roland A.; (2006): Dinosaur killer claws or climbing crampons? Biology Letters, 2 pp. 110-112.
|
|
|
|
Post by Infinity Blade on Oct 26, 2015 7:57:45 GMT 5
|
|
|
|
Post by Infinity Blade on Jul 9, 2016 23:16:43 GMT 5
I just found this publication about how quickly Tyrannosaurus' forelimbs could accelerate and thus how useful they could be in grabbing an escaping animal. It was concluded that they were quick enough to do so. I read an article reporting it a while ago, but I never found the actual publication until now. link |
|
|
|
Post by Infinity Blade on Aug 2, 2016 20:35:51 GMT 5
|
|
|
|
Post by Infinity Blade on Aug 25, 2016 7:16:38 GMT 5
Form and function of the foot and hallux in theropod dinosaursThe presence of an opposed hallux (digit I) in theropod lineage is regarded as an important indicator of increasing perching ability. However, the function of the foot with non-reversed hallux of non-avian theropods is poorly understood so far. To clarify the foot function of non-avian theropods, principal component (PC) analyses were conducted on four datasets based on linear measurement values on all non-ungual pedal phalanges in extant birds, and then, respective datasets of non-avian theropods were compared to them. One of these analyses successfully differentiated the plot distributions among extant birds in different functional categories of foot, and plots of non-avian theropod dinosaurs in the morphospace distributed in close to the convex hull of ground foraging bird plots. Additionally, detailed observation was done for the purpose of unraveling the hallucal functions. According to the form of articulation, the movement axis of hallucal metatarso-phalangeal joint in general theropods were more restricted to vertical motion than extant ground-foraging birds. This might indicate mechanical function of hallux, such as prey holding. In three specimens of Velociraptor (Dromaeosauridae ), attachment of MT I on the MT II varied from the medial to the plantar side, and which is respectively accompanied by the lateral and posterior directions of the distal articular facet of MT I and associated digits. Similar condition of MT I was seen in three troodontid specimens, and the proximal articular facets of MT I of both dromaeosaurid and troodontid specimens were convex like one of extant birds, rather than the concave facet of more basal theropods. These characters mean the mobility of intermetatarsal joint between MT I and II was increased in Paraves, and then the range of hallucal movement was extended, the feature presumably correlated with acquisition of the perch grasping function in basal birds.link
|
|
|
|
Post by Infinity Blade on Sept 5, 2016 19:36:50 GMT 5
Forearm Range of Motion in Australovenator wintonensis (Theropoda, Megaraptoridae)The hypertrophied manual claws and modified manus of megaraptoran theropods represent an unusual morphological adaptation among carnivorous dinosaurs. The skeleton of Australovenator wintonensis from the Cenomanian of Australia is among the most complete of any megaraptorid. It presents the opportunity to examine the range of motion of its forearm and the function of its highly modified manus. This provides the basis for behavioural inferences, and comparison with other Gondwanan theropod groups. Digital models created from computed tomography scans of the holotype reveal a humerus range of motion that is much greater than Allosaurus, Acrocanthosaurus, Tyrannosaurus but similar to that of the dromaeosaurid Bambiraptor. During flexion, the radius was forced distally by the radial condyle of the humerus. This movement is here suggested as a mechanism that forced a medial movement of the wrist. The antebrachium possessed a range of motion that was close to dromaeosaurids; however, the unguals were capable of hyper-extension, in particular manual phalanx I-2, which is a primitive range of motion characteristic seen in allosaurids and Dilophosaurus. During flexion, digits I and II slightly converge and diverge when extended which is accentuated by hyperextension of the digits in particular the unguals. We envision that prey was dispatched by its hands and feet with manual phalanx I-2 playing a dominant role. The range of motion analysis neither confirms nor refutes current phylogenetic hypotheses with regards to the placement of Megaraptoridae; however, we note Australovenator possessed, not only a similar forearm range of motion to some maniraptorans and basal coelurosaurs, but also similarities with Tetanurans (Allosauroids and Dilophosaurus). journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0137709The pes of Australovenator wintonensis (Theropoda: Megaraptoridae): analysis of the pedal range of motion and biological restorationThe ROM of pedal phalanx II-3 was obscured by a pathology on its articular facet. The bone is splayed outward as a thin lip, possibly resulting from constant impacts. What caused the pathology is unknown however we speculate that the ungual could have been used as a wounding implement much like the extant ratite Casuarius casuarius (Linnaeus, 1758) (commonly known as a cassowary). This bird, along with Dromaius, are known to kick during intraspecific fights or when threatened (Davies, 2002; Kofron, 1999; Kofron, 2003). One particular zookeeper described being wounded on two separate occasions by a sandhill crane and an emu. In both cases the impact was initiated with the second digit (Senter, 2009). Casuarius are renowned for kicking and interestingly, like Australovenator, the largest of the pedal unguals is on the second digit. Consequently, the latter might be a plausible explanation for the existence of the proximal pathology on the ungual.
Interestingly, the dissection of the Dromaius pes revealed the extensive mobility of the second toe compared to digits III and IV. This mobility was visualized with the manual manipulation of the FPDII which was not anchored to the metatarsus as were FPDIII and FPDIV. This increased mobility associated with the soft tissue structure and the behavioral aspect of kicking demonstrated in both Dromaius and Casuarius possibly explains the existence of the pathology on pedal phalanx II-3 of Australovenator.
peerj.com/articles/2312/
|
|
|
|
Post by Saurischian on Jan 5, 2017 19:42:23 GMT 5
Regarding Australovenator,this is what Matt A. White et al. says: "The forearm ROM comparisons of Australovenator with other theropod groups provide insight into the functional role the forelimb played during prey capture and dispatch. Larger theropods such as tyrannosaurids and carcharodontosaurids possessed limited anterior reach suggesting that initial prey capture was made orally with the forearms used to maintain grip on prey.The forearm lateral extension mobility of Australovenator possibly represented a predatory action that facilitated prey capture. The reasonably large ROM of the antebrachium indicates a large forearm extension and infers the capability to draw in the arms (and prey) close to the chest for easier dispatch. Australovenator had a gracile dentary and most likely possessed a relatively weak bite as a consequence. However, like tyrannosaurids, the unguals of digits I and II converged during flexion, which accentuated grip in an action of drawing the arms closer to the chest. There is also some minor mediolateral movement achieved between the proximal phalanx of digit I and metacarpal I. This movement was relatively restricted compared to the dromaeosaurid Bambiraptor, which ostensibly possessed an opposable digit I. As a result, Australovenator most likely required both hands to secure prey. Theropods with relatively weak bites relied on their hands and feet to assist in the dispatch of prey; however, the approach varied depending on the ROM and morphology of the forearms.
"
"Australovenator appears to be closer in manual ROM to Dilophosaurus than to any other theropod, whereby the first digit hypothetically played a dominant role in prey dispatch. Smaller theropods such as dromaeosaurids possessed greater manual flexion but lacked the extension capabilities of Australovenator. This emphasizes that the main function of the dromaeosaurid manus was to grapple, with dispatch achieved orally and/or with the sickle-like claw on its second pedal digit.Hyperextension of the manual unguals of Australoventor is greater than in any other theropod compared. Subsequently this infers that Australovenator and megaraptorids alike possessed a unique manus function most likely associated with predation."It appears that,although Australovenator(and probably other megaraptorans)had very impressive forearm range of motion,forelimb strength(as suggested by the well developed deltopectoral crest and olecranon process),enlarged claws likely with sharp ventral borders,and other adaptations that increase the ability of prey acquisition and dispacth with the forearms,the jaws are relatively gracile(at least going by the dentary).I think we see here something the opposite we see in animals like large carnosaurs,that probably used the jaws as main tools for prey acquisition and the clawed arms as secondary tools;here,the limbs seem to be the main tools.What would led Australovenator and kin to evolve so differently? --- Regarding noasaurid dinosaurs:Masiakasaurus,according to Carrano et al. (2011),had a number of adaptations on the shoulder bones and humeri that would likely give it an impressive forearm strength and range of motion.However,the manual claws doesn't look very well adapted for seizing other animals: 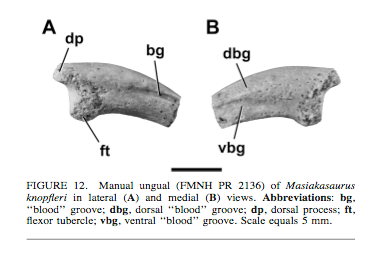 It looks blunt,in fact.This,and other adaptations,led to Duane Nash believe that it was using its limbs for digging,and that it was some type of dinosaurian ethiopian wolf,a specialist hunter of fossorial species.You can see it here: antediluviansalad.blogspot.com.br/2015/09/masiakasaurus-proposed-as-fossorial.htmlBy the other hand,the relative Noasaurus leali seems to have,like megaraptorans,large,strongly curved and trenchant claws;these claws were originally believed to be on the foot,but most recent analysis show that they were,again like megaraptorans,placed on the hand.  The placement of the claws on the hand would make sense,given the well development of the forearm in noasauridae.However,unlike megaraptorans,the jaw material of Noasaurus leali seems not to be gracile;in fact,it is said to be comparable to the jaw material of the larger abelisaurids(which are well known for having strong skulls). --- As for Therizinosaurus cheloniformis,I think this may be interesting: From Stephan Lautenschlager(2014): "While the short and compact manual ungual morphology of Alxasaurus recorded only very low von Mises stress, the stress magnitude increased with the curvature and particularly with the elongation of the unguals in the other taxa regardless of the tested functional scenario. Stress was most pronounced in the hypertrophied claw morphology of Therizinosaurus. The same holds true for displacement and strain magnitudes"
"Similarly, a comparison between the different functional scenarios reveals that the highest von Mises stress (as well as peak compressive and tensile stress, which coincide with von Mises peak stress), displacement and strain magnitudes were recorded for a scratch-digging function, in which the force is applied on the ventral tip of the claw. In this scenario, stress and strain hot-spots were focused along the dorsal surface of the claws and were again most pronounced in the elongate claws of Therizinosaurus, Nothronychus and Falcarius. By contrast, the hook-and-pull scenario resulted in lower magnitudes than scratch-digging, but with a similar pattern of stress distribution. The overall lowest magnitudes were found in the scenario simulating a piercing function with the force directed opposite the claw tip. In this scenario, stress and strain hot-spots occurred along the ventral surface of the tip with the rest of the claw largely unaffected."
"the more strongly curved and elongate unguals of the other sampled therizinosaurian taxa recorded the highest stress magnitudes when simulating a scratch-digging scenario, indicating this as the most unlikely of the tested functionalities."

"For ornithomimosaurs, which share a similar ungual morphology and morphospace occupation with several therizinosaurian taxa, such as Therizinosaurus and Beipiaosaurus, range-of-motion studies suggested the use of their manus in a hook-and-pull fashion to pull vegetation within reach. Following the results of the FEA and the shared morphospace occupation, a similar function is likely for therizinosaurs possessing elongate and curved unguals. However, if the unguals were used for browsing and pulling down vegetation, it would be expected that the forelimbs served to extend the range of the animal to a point, which cannot be reached by the head Therizinosaurian taxa, in which both cervical as well as forelimb elements are preserved, indicate that the neck is nearly equal in length or longer than the forelimbs (Nothronychus: neck = 118% of forelimbs, Alxasaurus: neck = 105% of forelimbs, Falcarius: neck = 103% of forelimbs). Thus, pulling of vegetation makes sense only, if large elements in reach were pulled down, in order to get at parts that were out of reach (e.g. long branches pulled down by a basal part)."
"Surprisingly, and despite their frequent portrayal as such in popular culture, there is no evidence that the hypertrophied claws of Therizinosaurus or other taxa would have been used in a defensive or aggressive context, although it seems plausible that the animal would make use of its claws when threatened. Similarly, the fragmentary fossil record for therizinosaurs makes it difficult to evaluate whether the enlarged unguals could represent a sexually dimorphic feature, as in some extant turtles"
The conclusion: Sources: -The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution;Federico L. Agnolin,Pablo Chiarelli. - New materials of Masiakasaurus knopfleri Sampson, Carrano, and Forster, 2001, and implications for the morphology of the Noasauridae (Theropoda: Ceratosauria);Carrano, M.T.; Loewen, M.A.; Sertic, J.J.W. -Forearm Range of Motion in Australovenator wintonensis (Theropoda, Megaraptoridae);Matt A. White , Phil R. Bell, Alex G. Cook, David G. Barnes, Travis R. Tischler, Brant J. Bassam, David A. Elliott. -Morphological and functional diversity in therizinosaur claws and the implications for theropod claw evolution;Stephan Lautenschlager. |
|
|
|
Post by Infinity Blade on Jan 6, 2017 2:29:07 GMT 5
Regarding Australovenator,this is what Matt A. White et al. says: "The forearm ROM comparisons of Australovenator with other theropod groups provide insight into the functional role the forelimb played during prey capture and dispatch. Larger theropods such as tyrannosaurids and carcharodontosaurids possessed limited anterior reach suggesting that initial prey capture was made orally with the forearms used to maintain grip on prey.The forearm lateral extension mobility of Australovenator possibly represented a predatory action that facilitated prey capture. The reasonably large ROM of the antebrachium indicates a large forearm extension and infers the capability to draw in the arms (and prey) close to the chest for easier dispatch. Australovenator had a gracile dentary and most likely possessed a relatively weak bite as a consequence. However, like tyrannosaurids, the unguals of digits I and II converged during flexion, which accentuated grip in an action of drawing the arms closer to the chest. There is also some minor mediolateral movement achieved between the proximal phalanx of digit I and metacarpal I. This movement was relatively restricted compared to the dromaeosaurid Bambiraptor, which ostensibly possessed an opposable digit I. As a result, Australovenator most likely required both hands to secure prey. Theropods with relatively weak bites relied on their hands and feet to assist in the dispatch of prey; however, the approach varied depending on the ROM and morphology of the forearms.
"
"Australovenator appears to be closer in manual ROM to Dilophosaurus than to any other theropod, whereby the first digit hypothetically played a dominant role in prey dispatch. Smaller theropods such as dromaeosaurids possessed greater manual flexion but lacked the extension capabilities of Australovenator. This emphasizes that the main function of the dromaeosaurid manus was to grapple, with dispatch achieved orally and/or with the sickle-like claw on its second pedal digit.Hyperextension of the manual unguals of Australoventor is greater than in any other theropod compared. Subsequently this infers that Australovenator and megaraptorids alike possessed a unique manus function most likely associated with predation."I already posted the abstract of this paper. The tip looks like it was broken off, likely through taphonomy. |
|
|
|
Post by Infinity Blade on Feb 8, 2017 8:01:14 GMT 5
|
|
|
|
Post by Infinity Blade on Feb 26, 2017 0:15:34 GMT 5
I noticed that the ventrolateral(?) margins of eagle talons are kind of keeled. 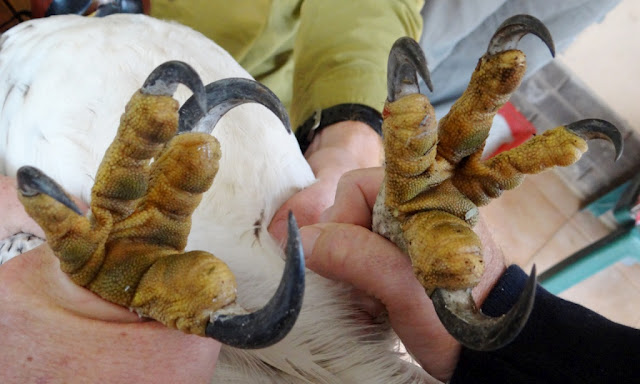 Does anyone think these keels assist in piercing (and secondarily cutting) prey? I know cat claws, which have some kind of a cutting edge, perform this function. www.ncbi.nlm.nih.gov/pmc/articles/PMC2736126/#!po=48.3696 And what about Mesozoic theropods? |
|
|
|
Post by Infinity Blade on Mar 19, 2017 18:30:20 GMT 5
journals.plos.org/plosone/article?id=10.1371/journal.pone.0028964#s4
I also wanted to know. Theropods already tended to have really large leg and hip muscles for locomotion, which would also make kicking quite powerful. But they also had some (huge) tail muscles (like the M. caudofemoralis), which also played a role in locomotion. Wouldn't they also amplilfy kicking power (possibly beyond what we see in ungulates or ratites)?
|
|












