


Post by Infinity Blade on Mar 26, 2015 6:32:48 GMT 5
Teleoceras spp.

© @ Eurwentala
An illustration of T. major next to H. amphibius for comparison.
Temporal range: Neogene; Miocene to Pliocene; late Hemingfordian to late Hemphillian (~17.5-4.9 Ma)[1]
Scientific classification:
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Superphylum: Deuterostomia
Phylum: Chordata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Clade: Synapsida
Clade: Eupelycosauria
Clade: Sphenacodontia
Clade: Sphenacodontoidea
Order: Therapsida
Suborder: Cynodontia
Clade: Prozostrodontia
Clade: Mammaliaformes
Class: Mammalia
Legion: Cladotheria
Sublegion: Zatheria
Infralegion: Tribosphenida
Subclass: Theria
Clade: Eutheria
Infraclass: Placentalia
Subcohort: Exafroplacentalia
Magnorder: Boreoeutheria
Superorder: Laurasiatheria
Clade: Ungulata
Order: Perissodactyla
Suborder: Ceratomorpha
Superfamily: Rhinocerotoidea
Family: Rhinocerotidae
Tribe: †Teleoceratini
Genus: †Teleoceras
Species: †T. aepysoma[2]
†T. aginense
†T. americanum
†T. brachyrhinum
†T. fossiger
†T. guymonense
†T. hicksi
†T. major
†T. medicornutum
†T. meridianum
†T. proterum
Teleoceras is an extinct genus of rhinoceros that lived in North America from the Miocene to Pliocene epochs (late Hemingfordian to late Hemphillian NALMAs). There were multiple species in this genus.
Evolution:
It has been proposed that Teleoceras evolved from Diaceratherium, as the latter genus spread to North America. According to Donald Prothero, the early species were themselves quite short-legged. The limb bones became stumpier and more compressed to an extreme degree as the genus evolved.[3] However, the discovery of T. aepysoma remains from the late Hemphillian[2] suggests that this evolutionary trend was not followed by all lineages in the genus Teleoceras.
Description:
Most Teleoceras species possessed a small nasal horn, fused nasal bones, and a hippopotamus-like body consisting of a barrel-shaped trunk and short, robust limbs.[2][3] There are many other synapomorphies of Teleoceras proposed by Donald Prothero, which T. aepysoma shares with its relatives. This has led to its inclusion in the genus. However, this particular species lacked the small terminal nasal horn, barrel-shaped trunk, and stumpy limbs of other Teleoceras species. The presence of a nasal incision retracted to the anterior third premolar, also thought to be a synapomorphy of the genus, was questionable in T. aepysoma. Accordingly, the synapomorphies of Teleoceras have been revised, with the specifically aforementioned ones being removed.[2]
Teleoceras possessed upper chisel-like incisors (unlike most aceratheriines).[3] Some specimens possess large, tusk-like second lower incisors, while in others they are smaller. Specimens with tusk-like i2s were male, while those with smaller incisors were female.[2] The skull was very large, with high-crowned molars that were almost certainly used to feed on abrasive grasses.[3]
Two male specimens of T. aepysoma were estimated to have weighed 1,544 kg and 2,268 kg, respectively. This is substantially heavier than mass estimates of 568-702 kg for T. proterum.[2]
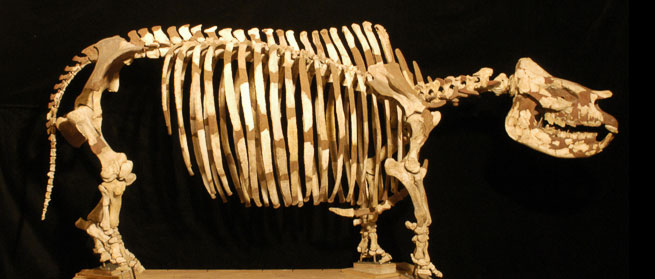
Mounted skeleton of T. proterum at the Florida Museum of Natural History.

A mounted skeleton of Teleoceras at the Harvard Museum of Natural History (image source).

Mounted cast of T. aepysoma at the East Tennessee State University Museum of Natural History, Gray, Tennessee.[2] Note the proportionately longer limbs in comparison to other species.
Paleobiology:
Because of its hippo-like proportions, Teleoceras was once thought to have led a hippo-like semiaquatic lifestyle.[3] However, oxygen isotope analysis of Teleoceras tooth enamel is inconsistent with this interpretation. δ18O values of a semiaquatic hippo-ecomorph would be expected to be much lower than those of contemporaneous terrestrial animals. Those of Teleoceras are actually similar to other Ashfall mammals.[1][4]
δ13C values show that for the majority of its history, Teleoceras was primarily consuming C3 plants. These would likely have been browse and C3 grasses. There is evidence that as C4 vegetation expanded (along with cooling global temperatures), Teleoceras was able to incorporate a modest amount of C4 plants in its diet[1][4], but it and other rhinoceroses were never as successful at doing so as contemporaneous horses.[4]
Fossil assemblages of Teleoceras are dominated by subadult and young adult male individuals that were between 15-40% of their potential lifespan. The age-specific mortality rates were virtually identical to those of modern black rhinoceros populations. Modern rhinos are characterized by a high rate of intraspecific fight-related mortality, strongly suggesting that this accounted for the high mortality rate seen in Teleoceras. This is in contrast to mortality rates in hippos (which, as noted above, Teleoceras has been compared to), which are not elevated.[5] Pronounced intermale aggression is also supported by the delayed eruption of i2 in male Teloceras, as well as the tusk facets that were continually sharpened by the upper incisors. However, body size dimorphism varies within each assemblage, suggesting that the intensity of intermale competition is unrelated to the degree of sexual dimorphism.[6]
In Nebraska, a fossil assemblage (now colloquially known as “Rhino Pompeii”) of hundreds of T. major individuals was uncovered. Out of more than 200 individuals collected in the first few years, only 7 were adult males; the rest were females with their calves (some of which even had their calves in a nursing position under their bellies, while others had fetuses in their pelvic cavities). This shows that Teleoceras lived in large, male-dominated herds composed mostly of females and their young, just like many large ungulates today. Likewise, these rhinos were buried under ash blown from the Rocky Mountains. In addition to those that slowly died or were suffocated after being buried alive, bone pathologies indicate that many died by inhaling razor-sharp volcanic ash shards (which would have lacerated their lungs). Lastly, some of the specimens had seeds of Berriochloa (a common grass from the time period) in close proximity with their throat bones, providing further evidence that Teleoceras grazed.[3]
Extinction:
Teleoceras went extinct during the late Hemphillian about 4.9 million years ago.[1] As the global climate cooled, C4 plants expanded and primary productivity would have declined. Rhinoceros abundance rapidly declined about 6.5 million years ago, coinciding with this expansion. As noted, while Teleoceras was able to incorporate C4 plants in its diet to a certain extent, rhinos were not as successful at doing this as contemporaneous horses. Therefore, Teleoceras likely went extinct as a result of declining food resources due to the expansion of C4 vegetation.[1]
References:
[1] Wang, B., & Secord, R. (2020). Paleoecology of Aphelops and Teleoceras (Rhinocerotidae) through an interval of changing climate and vegetation in the Neogene of the Great Plains, central United States. Palaeogeography, Palaeoclimatology, Palaeoecology, 542, 109411.
[2] Short, R. A., Wallace, S. C., & Emmert, L. G. (2019). A new species of Teleoceras (Mammalia, Rhinocerotidae) from the late Hemphillian of Tennessee. Bull Florida Mus Nat Hist, 56, 183-260.
[3] Prothero, D. R., & Schoch, R. M. (2002). Horns, tusks, and flippers: the evolution of hoofed mammals. JHU Press.
[4] MacFadden, B. J. (1998). [url=https://www.jstor.org/stable/2401243?seq=1#metadata_info_tab_contentsTale of two rhinos: isotopic ecology, paleodiet, and niche differentiation of Aphelops and Teloceras from the Florida Neogene[/url]. Paleobiology, 274-286.
[5] Mihlbachler, M. C. (2003). Demography of late Miocene rhinoceroses (Teleoceras proterum and Aphelops malacorhinus) from Florida: linking mortality and sociality in fossil assemblages. Paleobiology, 29(3), 412-428.
[6] Mihlbachlerll, M. C. (2005). LINKING SEXUAL DIMORPHISMAND SOCIALITY IN RHINOCEROSES: INSIGHTS FROM TELEOCERAS PROTER UM AND APHELOPS AMLA CORHIN US FROM THE LATE MIOCENE OF FLORIDA. Bull. Fla. Mus. Nat. Hist, 45(4), 495-520.
© @ Eurwentala
An illustration of T. major next to H. amphibius for comparison.
Temporal range: Neogene; Miocene to Pliocene; late Hemingfordian to late Hemphillian (~17.5-4.9 Ma)[1]
Scientific classification:
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Superphylum: Deuterostomia
Phylum: Chordata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Clade: Synapsida
Clade: Eupelycosauria
Clade: Sphenacodontia
Clade: Sphenacodontoidea
Order: Therapsida
Suborder: Cynodontia
Clade: Prozostrodontia
Clade: Mammaliaformes
Class: Mammalia
Legion: Cladotheria
Sublegion: Zatheria
Infralegion: Tribosphenida
Subclass: Theria
Clade: Eutheria
Infraclass: Placentalia
Subcohort: Exafroplacentalia
Magnorder: Boreoeutheria
Superorder: Laurasiatheria
Clade: Ungulata
Order: Perissodactyla
Suborder: Ceratomorpha
Superfamily: Rhinocerotoidea
Family: Rhinocerotidae
Tribe: †Teleoceratini
Genus: †Teleoceras
Species: †T. aepysoma[2]
†T. aginense
†T. americanum
†T. brachyrhinum
†T. fossiger
†T. guymonense
†T. hicksi
†T. major
†T. medicornutum
†T. meridianum
†T. proterum
Teleoceras is an extinct genus of rhinoceros that lived in North America from the Miocene to Pliocene epochs (late Hemingfordian to late Hemphillian NALMAs). There were multiple species in this genus.
Evolution:
It has been proposed that Teleoceras evolved from Diaceratherium, as the latter genus spread to North America. According to Donald Prothero, the early species were themselves quite short-legged. The limb bones became stumpier and more compressed to an extreme degree as the genus evolved.[3] However, the discovery of T. aepysoma remains from the late Hemphillian[2] suggests that this evolutionary trend was not followed by all lineages in the genus Teleoceras.
Description:
Most Teleoceras species possessed a small nasal horn, fused nasal bones, and a hippopotamus-like body consisting of a barrel-shaped trunk and short, robust limbs.[2][3] There are many other synapomorphies of Teleoceras proposed by Donald Prothero, which T. aepysoma shares with its relatives. This has led to its inclusion in the genus. However, this particular species lacked the small terminal nasal horn, barrel-shaped trunk, and stumpy limbs of other Teleoceras species. The presence of a nasal incision retracted to the anterior third premolar, also thought to be a synapomorphy of the genus, was questionable in T. aepysoma. Accordingly, the synapomorphies of Teleoceras have been revised, with the specifically aforementioned ones being removed.[2]
Teleoceras possessed upper chisel-like incisors (unlike most aceratheriines).[3] Some specimens possess large, tusk-like second lower incisors, while in others they are smaller. Specimens with tusk-like i2s were male, while those with smaller incisors were female.[2] The skull was very large, with high-crowned molars that were almost certainly used to feed on abrasive grasses.[3]
Two male specimens of T. aepysoma were estimated to have weighed 1,544 kg and 2,268 kg, respectively. This is substantially heavier than mass estimates of 568-702 kg for T. proterum.[2]
Mounted skeleton of T. proterum at the Florida Museum of Natural History.
A mounted skeleton of Teleoceras at the Harvard Museum of Natural History (image source).
Mounted cast of T. aepysoma at the East Tennessee State University Museum of Natural History, Gray, Tennessee.[2] Note the proportionately longer limbs in comparison to other species.
Paleobiology:
Because of its hippo-like proportions, Teleoceras was once thought to have led a hippo-like semiaquatic lifestyle.[3] However, oxygen isotope analysis of Teleoceras tooth enamel is inconsistent with this interpretation. δ18O values of a semiaquatic hippo-ecomorph would be expected to be much lower than those of contemporaneous terrestrial animals. Those of Teleoceras are actually similar to other Ashfall mammals.[1][4]
δ13C values show that for the majority of its history, Teleoceras was primarily consuming C3 plants. These would likely have been browse and C3 grasses. There is evidence that as C4 vegetation expanded (along with cooling global temperatures), Teleoceras was able to incorporate a modest amount of C4 plants in its diet[1][4], but it and other rhinoceroses were never as successful at doing so as contemporaneous horses.[4]
Fossil assemblages of Teleoceras are dominated by subadult and young adult male individuals that were between 15-40% of their potential lifespan. The age-specific mortality rates were virtually identical to those of modern black rhinoceros populations. Modern rhinos are characterized by a high rate of intraspecific fight-related mortality, strongly suggesting that this accounted for the high mortality rate seen in Teleoceras. This is in contrast to mortality rates in hippos (which, as noted above, Teleoceras has been compared to), which are not elevated.[5] Pronounced intermale aggression is also supported by the delayed eruption of i2 in male Teloceras, as well as the tusk facets that were continually sharpened by the upper incisors. However, body size dimorphism varies within each assemblage, suggesting that the intensity of intermale competition is unrelated to the degree of sexual dimorphism.[6]
In Nebraska, a fossil assemblage (now colloquially known as “Rhino Pompeii”) of hundreds of T. major individuals was uncovered. Out of more than 200 individuals collected in the first few years, only 7 were adult males; the rest were females with their calves (some of which even had their calves in a nursing position under their bellies, while others had fetuses in their pelvic cavities). This shows that Teleoceras lived in large, male-dominated herds composed mostly of females and their young, just like many large ungulates today. Likewise, these rhinos were buried under ash blown from the Rocky Mountains. In addition to those that slowly died or were suffocated after being buried alive, bone pathologies indicate that many died by inhaling razor-sharp volcanic ash shards (which would have lacerated their lungs). Lastly, some of the specimens had seeds of Berriochloa (a common grass from the time period) in close proximity with their throat bones, providing further evidence that Teleoceras grazed.[3]
Extinction:
Teleoceras went extinct during the late Hemphillian about 4.9 million years ago.[1] As the global climate cooled, C4 plants expanded and primary productivity would have declined. Rhinoceros abundance rapidly declined about 6.5 million years ago, coinciding with this expansion. As noted, while Teleoceras was able to incorporate C4 plants in its diet to a certain extent, rhinos were not as successful at doing this as contemporaneous horses. Therefore, Teleoceras likely went extinct as a result of declining food resources due to the expansion of C4 vegetation.[1]
References:
[1] Wang, B., & Secord, R. (2020). Paleoecology of Aphelops and Teleoceras (Rhinocerotidae) through an interval of changing climate and vegetation in the Neogene of the Great Plains, central United States. Palaeogeography, Palaeoclimatology, Palaeoecology, 542, 109411.
[2] Short, R. A., Wallace, S. C., & Emmert, L. G. (2019). A new species of Teleoceras (Mammalia, Rhinocerotidae) from the late Hemphillian of Tennessee. Bull Florida Mus Nat Hist, 56, 183-260.
[3] Prothero, D. R., & Schoch, R. M. (2002). Horns, tusks, and flippers: the evolution of hoofed mammals. JHU Press.
[4] MacFadden, B. J. (1998). [url=https://www.jstor.org/stable/2401243?seq=1#metadata_info_tab_contentsTale of two rhinos: isotopic ecology, paleodiet, and niche differentiation of Aphelops and Teloceras from the Florida Neogene[/url]. Paleobiology, 274-286.
[5] Mihlbachler, M. C. (2003). Demography of late Miocene rhinoceroses (Teleoceras proterum and Aphelops malacorhinus) from Florida: linking mortality and sociality in fossil assemblages. Paleobiology, 29(3), 412-428.
[6] Mihlbachlerll, M. C. (2005). LINKING SEXUAL DIMORPHISMAND SOCIALITY IN RHINOCEROSES: INSIGHTS FROM TELEOCERAS PROTER UM AND APHELOPS AMLA CORHIN US FROM THE LATE MIOCENE OF FLORIDA. Bull. Fla. Mus. Nat. Hist, 45(4), 495-520.
