Post by dinosauria101 on Aug 30, 2019 16:27:29 GMT 5
Triceratops spp.

Image by Andrey Atuchin->
Temporal range: Late Cretaceous; Maastrichtian (68-66 Ma)
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Olfactores
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Clade: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Clade: Reptilia
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Clade: †Ornithischia
Clade: †Genasauria
Clade: †Neornithischia
Clade: †Cerapoda
Clade: †Marginocephalia
Clade: †Ceratopsia
Clade: †Neoceratopsia
Clade: †Coronosauria
Superfamily: †Ceratopsoidea
Family: †Ceratopsidae
Clade: †Chasmosaurinae
Tribe: †Triceratopsini
Genus: †Triceratops
Species: †T. horridus, †T. prorsus
Triceratops is a genus of ceratopsian dinosaur that lived in North America from 68-66 million years ago. It was one of the last non-avian dinosaur genera that ever lived, having gone extinct during the Cretaceous-Paleogene extinction event.
Description:
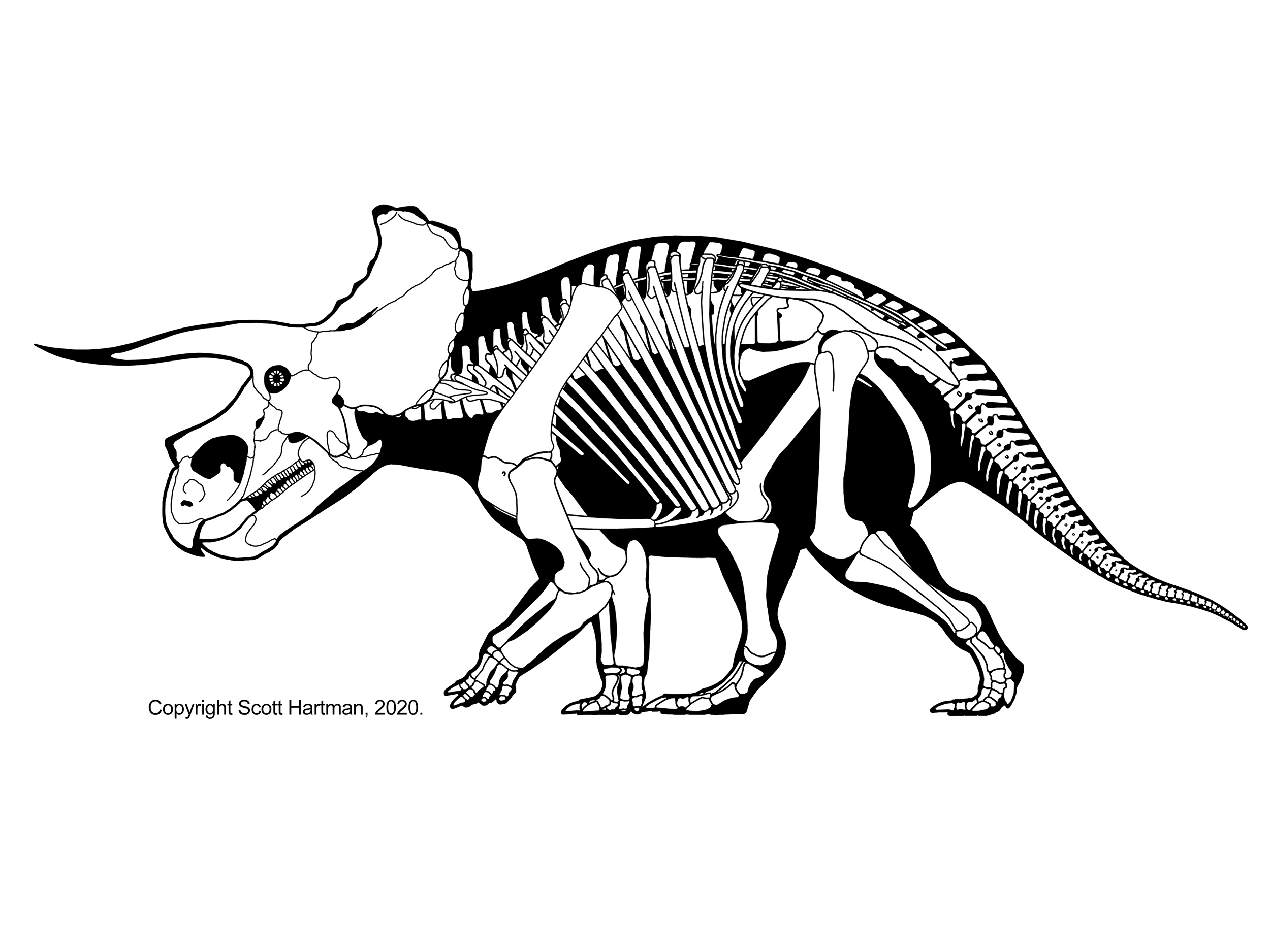
Triceratops was a large animal. Although body mass estimates vary, a handful seem to be more or less consistent with an animal weighing as much as a modern day African bush elephant, somewhere between ~6-9 tonnes.[1][2][3] The skeleton was very robust.[3]
There has been some debate as to whether or not the forelimbs of Triceratops and its kind were sprawled laterally or held erect. In more recent years, a 2012 analysis of elbow joint adductor moment arms in extinct quadrupedal tetrapods supports an upright/sagittal forelimb posture and function for Triceratops, and probably for large ceratopsids as a whole.[4] Unlike the African bush elephant, the only modern terrestrial quadruped in Triceratops’ size range, Triceratops’ limbs were not held in a columnar, elephantine stance, being flexed and flexible at the shoulder, elbow, knee, and ankle.[5][6][7] Such flexure and flexibility is necessary for the joints to act as active springs to propel an animal forward during relatively fast locomotion.[6] This, coupled with other appendicular adaptations possessed by ceratopsids like Triceratops (namely proportionately large attachment points and areas for leg musculature[3][7][8][9], as well as high limb bone strength[6][7][9][10]) suggest that, while Triceratops and kin were not highly cursorial animals, their locomotory performance would have exceeded that of other giant, but fully graviportal animals (e.g. elephants).[10]
The name Triceratops literally translates to “three-horned face”, in reference to the three horns of the genus. Along with these horns was a thickened solid frill of bone at the back of the skull. Their function has been the subject of study over the years. That species recognition was the driver of the diversity of ceratopsid cranial ornamentation is not well supported.[11][12] The two main hypotheses are that the cranial ornaments of ceratopsids (Triceratops included) evolved for sexual selection and for use as weapons/defense.[12] Evidence for head-to-head encounters with the giant predator Tyrannosaurus rex involving horns, and intraspecific combat, both exist.[13][14] The horn cores of Triceratops were as thick relative to body mass as the tusks of proboscideans like the African bush elephant, mammoths, and the American mastodon (namely the basal areas thereof). Unlike elephant tusks, however, the horn cores of Triceratops were further strengthened by a keratinous sheath, and the material properties plus the amount of interior hollow space for both were different. A plot between horn/tusk basal area relative to length (or reach) shows that, while proboscidean tusks continue the trend seen in antelope horns, Triceratops and other ceratopsian brow horns plot well above this trend. This means that the horn cores of Triceratops were relatively short and thick, likely making them better adapted to withstand forces distal to their bases than the horns of antelope and the tusks of proboscideans.[15] The frill of Triceratops was composed of fibrolamellar bone[16], which is critical in wound healing.[17][18]

Graph plotting horn/tusk basal area against length/reach. Note the tendency of ceratopsid orbital horn cores to plot above the horns of antelope and the tusks of proboscideans. Screen capture from [15].
The head also sported a toothless hooked beak. Triceratops and other ceratopsids exhibited precise dental occlusion in their high-angled slicing teeth.[19] The teeth grew in batteries, where 36-40 tooth columns, each stacked with 3-5 teeth, existed on each side of each jaw. In total, anywhere from 432 to 800 teeth could exist in an individual Triceratops’ mouth, but only a portion of these teeth were used for feeding at any given time.[20] The tooth crowns of ceratopsians were complex, especially in protoceratopsids and ceratopsids. The teeth of Triceratops had five layers of wear-relevant tissue: enamel, hard mantle dentine, orthodentine, vasodentine, and coronal cementum. This exceeds the amount of the tooth tissue present in the most complex mammalian dentitions (like those of ungulates), which possess four layers of tissue (all of the aforementioned tissues minus vasodentine), and greatly exceeds the number of tooth tissues typically seen in reptiles (enamel and orthodentine).[19] Only hadrosaurids are known to have evolved more complex teeth, which had six layers.[21] The vasodentine in particular was very important. Because vasodentine is exceptionally porous, it was worn frequently to form fullers on the chewing surfaces of the teeth. This unique morphology reduced friction while processing food[19], and therefore would have given Triceratops teeth that rivaled, if not exceeded, the teeth of modern herbivorous mammals in complexity.[19][21]

Triceratops tooth crown histology. Fig. 2 of [19].
Skin impressions from Triceratops suggest a scale morphology markedly different from those of other ceratopsians. These scales tended to be larger than those of other ceratopsids, with the largest having raised cones/tubercles that were widely spaced in an irregular pattern.[3][22]

Skin impressions from a Wyoming Triceratops known as “Lane”. Taken from [22].
Ontogeny:
Triceratops underwent major ontogenetic changes. These include, but are not limited to, the margin of the frill transitioning from scalloped to wavy and smooth, internal hollowing of the base of the postorbital horns, the orbital horns transitioning from curving backwards to pointing forwards, etc.[23]
References:
[1] Campione, N. E., & Evans, D. C. (2012). A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. Bmc Biology, 10(1), 60.
[2] Alexander, R. M. (1985). Mechanics of posture and gait of some large dinosaurs. Zoological journal of the linnean society, 83(1), 1-25.
[3] Paul, G. S. (2016). The Princeton field guide to dinosaurs (Vol. 110). Princeton University Press. pp. 301-303.
[4] Fujiwara, S. I., & Hutchinson, J. R. (2012). Elbow joint adductor moment arm as an indicator of forelimb posture in extinct quadrupedal tetrapods. Proceedings of the Royal Society B: Biological Sciences, 279(1738), 2561-2570.
[5] Paul, G. S., & Christiansen, P. (2000). Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion. Paleobiology, 26(3), 450-465.
[6] Paul, G. S. (1998). Limb design, function and running performance in ostrich-mimmics and tyrannosaurs. GAIA: revista de geociências, (15), 257.
[7] Paul, G. (2003). The Scientific American book of dinosaurs. Macmillan. p. 74
[8] Brett-Surman, M. K., Holtz, T. R., & Farlow, J. O. (Eds.). (2012). The complete dinosaur. Indiana University Press. p. 865.
[9] Bakker, R. T. (1986). The dinosaur heresies. William Morrow.
[10] Christiansen, P., & Paul, G. S. (2001). Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs. Gaia, 16, 13-29.
[11] Hone, D. W. E., & Naish, D. (2013). The ‘species recognition hypothesis’ does not explain the presence and evolution of exaggerated structures in non‐avialan dinosaurs. Journal of Zoology, 290(3), 172-180.
[12] Knapp, A., Knell, R. J., Farke, A. A., Loewen, M. A., & Hone, D. W. (2018). Patterns of divergence in the morphology of ceratopsian dinosaurs: sympatry is not a driver of ornament evolution . Proceedings of the Royal Society B: Biological Sciences, 285(1875), 20180312.
[13] Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. Tyrannosaurus rex the Tyrant king, 355-370.
[14] Farke, A. A., Wolff, E. D., & Tanke, D. H. (2009). Evidence of combat in Triceratops. PLoS One, 4(1), e4252.
[15] Farlow, J. O. (1990). Dynamic Dinosaurs-Dynamics of Dinosaurs and Other Extinct Giants. R. McNeill Alexander. Columbia University Press; New York. 1989. viii+ 167 pp. $30.00. Paleobiology, 16(2), 234-241.
[16] Reid, R.E.H. (1997). "Histology of bones and teeth". In Currie, P. J.; Padian, K. (eds.). Encyclopedia of Dinosaurs. San Diego, CA.: Academic Press. pp. 329–339.
[17] Horner, J. R., & Goodwin, M. B. (2009). Extreme cranial ontogeny in the Upper Cretaceous dinosaur Pachycephalosaurus. PLoS One, 4(10), e7626.
[18] Horner, J. R., & Lamm, E. T. (2011). Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis. Comptes Rendus Palevol, 10(5-6), 439-452.
[19] Erickson, G. M., Sidebottom, M. A., Kay, D. I., Turner, K. T., Ip, N., Norell, M. A., ... & Krick, B. A. (2015). Wear biomechanics in the slicing dentition of the giant horned dinosaur Triceratops. Science Advances, 1(5), e1500055.
[20] Dodson, P.; Forster, C. A.; Sampson, S. D. (2004). "Ceratopsidae". In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (second ed.). Berkeley: University of California Press. pp. 494–513. ISBN 978-0-520-24209-8.
[21] Erickson, G. M., Krick, B. A., Hamilton, M., Bourne, G. R., Norell, M. A., Lilleodden, E., & Sawyer, W. G. (2012). Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science, 338(6103), 98-101.
[22] markwitton-com.blogspot.com/2015/12/dinosaur-scales-some-thoughts-for.html
[23] Horner, J. R., & Goodwin, M. B. (2006). Major cranial changes during Triceratops ontogeny. Proceedings of the Royal Society B: Biological Sciences, 273(1602), 2757-2761.
Image by Andrey Atuchin->
Temporal range: Late Cretaceous; Maastrichtian (68-66 Ma)
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Olfactores
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Clade: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Clade: Reptilia
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Clade: †Ornithischia
Clade: †Genasauria
Clade: †Neornithischia
Clade: †Cerapoda
Clade: †Marginocephalia
Clade: †Ceratopsia
Clade: †Neoceratopsia
Clade: †Coronosauria
Superfamily: †Ceratopsoidea
Family: †Ceratopsidae
Clade: †Chasmosaurinae
Tribe: †Triceratopsini
Genus: †Triceratops
Species: †T. horridus, †T. prorsus
Triceratops is a genus of ceratopsian dinosaur that lived in North America from 68-66 million years ago. It was one of the last non-avian dinosaur genera that ever lived, having gone extinct during the Cretaceous-Paleogene extinction event.
Description:
Triceratops was a large animal. Although body mass estimates vary, a handful seem to be more or less consistent with an animal weighing as much as a modern day African bush elephant, somewhere between ~6-9 tonnes.[1][2][3] The skeleton was very robust.[3]
There has been some debate as to whether or not the forelimbs of Triceratops and its kind were sprawled laterally or held erect. In more recent years, a 2012 analysis of elbow joint adductor moment arms in extinct quadrupedal tetrapods supports an upright/sagittal forelimb posture and function for Triceratops, and probably for large ceratopsids as a whole.[4] Unlike the African bush elephant, the only modern terrestrial quadruped in Triceratops’ size range, Triceratops’ limbs were not held in a columnar, elephantine stance, being flexed and flexible at the shoulder, elbow, knee, and ankle.[5][6][7] Such flexure and flexibility is necessary for the joints to act as active springs to propel an animal forward during relatively fast locomotion.[6] This, coupled with other appendicular adaptations possessed by ceratopsids like Triceratops (namely proportionately large attachment points and areas for leg musculature[3][7][8][9], as well as high limb bone strength[6][7][9][10]) suggest that, while Triceratops and kin were not highly cursorial animals, their locomotory performance would have exceeded that of other giant, but fully graviportal animals (e.g. elephants).[10]
The name Triceratops literally translates to “three-horned face”, in reference to the three horns of the genus. Along with these horns was a thickened solid frill of bone at the back of the skull. Their function has been the subject of study over the years. That species recognition was the driver of the diversity of ceratopsid cranial ornamentation is not well supported.[11][12] The two main hypotheses are that the cranial ornaments of ceratopsids (Triceratops included) evolved for sexual selection and for use as weapons/defense.[12] Evidence for head-to-head encounters with the giant predator Tyrannosaurus rex involving horns, and intraspecific combat, both exist.[13][14] The horn cores of Triceratops were as thick relative to body mass as the tusks of proboscideans like the African bush elephant, mammoths, and the American mastodon (namely the basal areas thereof). Unlike elephant tusks, however, the horn cores of Triceratops were further strengthened by a keratinous sheath, and the material properties plus the amount of interior hollow space for both were different. A plot between horn/tusk basal area relative to length (or reach) shows that, while proboscidean tusks continue the trend seen in antelope horns, Triceratops and other ceratopsian brow horns plot well above this trend. This means that the horn cores of Triceratops were relatively short and thick, likely making them better adapted to withstand forces distal to their bases than the horns of antelope and the tusks of proboscideans.[15] The frill of Triceratops was composed of fibrolamellar bone[16], which is critical in wound healing.[17][18]
Graph plotting horn/tusk basal area against length/reach. Note the tendency of ceratopsid orbital horn cores to plot above the horns of antelope and the tusks of proboscideans. Screen capture from [15].
The head also sported a toothless hooked beak. Triceratops and other ceratopsids exhibited precise dental occlusion in their high-angled slicing teeth.[19] The teeth grew in batteries, where 36-40 tooth columns, each stacked with 3-5 teeth, existed on each side of each jaw. In total, anywhere from 432 to 800 teeth could exist in an individual Triceratops’ mouth, but only a portion of these teeth were used for feeding at any given time.[20] The tooth crowns of ceratopsians were complex, especially in protoceratopsids and ceratopsids. The teeth of Triceratops had five layers of wear-relevant tissue: enamel, hard mantle dentine, orthodentine, vasodentine, and coronal cementum. This exceeds the amount of the tooth tissue present in the most complex mammalian dentitions (like those of ungulates), which possess four layers of tissue (all of the aforementioned tissues minus vasodentine), and greatly exceeds the number of tooth tissues typically seen in reptiles (enamel and orthodentine).[19] Only hadrosaurids are known to have evolved more complex teeth, which had six layers.[21] The vasodentine in particular was very important. Because vasodentine is exceptionally porous, it was worn frequently to form fullers on the chewing surfaces of the teeth. This unique morphology reduced friction while processing food[19], and therefore would have given Triceratops teeth that rivaled, if not exceeded, the teeth of modern herbivorous mammals in complexity.[19][21]
Triceratops tooth crown histology. Fig. 2 of [19].
Skin impressions from Triceratops suggest a scale morphology markedly different from those of other ceratopsians. These scales tended to be larger than those of other ceratopsids, with the largest having raised cones/tubercles that were widely spaced in an irregular pattern.[3][22]
Skin impressions from a Wyoming Triceratops known as “Lane”. Taken from [22].
Ontogeny:
Triceratops underwent major ontogenetic changes. These include, but are not limited to, the margin of the frill transitioning from scalloped to wavy and smooth, internal hollowing of the base of the postorbital horns, the orbital horns transitioning from curving backwards to pointing forwards, etc.[23]
References:
[1] Campione, N. E., & Evans, D. C. (2012). A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. Bmc Biology, 10(1), 60.
[2] Alexander, R. M. (1985). Mechanics of posture and gait of some large dinosaurs. Zoological journal of the linnean society, 83(1), 1-25.
[3] Paul, G. S. (2016). The Princeton field guide to dinosaurs (Vol. 110). Princeton University Press. pp. 301-303.
[4] Fujiwara, S. I., & Hutchinson, J. R. (2012). Elbow joint adductor moment arm as an indicator of forelimb posture in extinct quadrupedal tetrapods. Proceedings of the Royal Society B: Biological Sciences, 279(1738), 2561-2570.
[5] Paul, G. S., & Christiansen, P. (2000). Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion. Paleobiology, 26(3), 450-465.
[6] Paul, G. S. (1998). Limb design, function and running performance in ostrich-mimmics and tyrannosaurs. GAIA: revista de geociências, (15), 257.
[7] Paul, G. (2003). The Scientific American book of dinosaurs. Macmillan. p. 74
[8] Brett-Surman, M. K., Holtz, T. R., & Farlow, J. O. (Eds.). (2012). The complete dinosaur. Indiana University Press. p. 865.
[9] Bakker, R. T. (1986). The dinosaur heresies. William Morrow.
[10] Christiansen, P., & Paul, G. S. (2001). Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs. Gaia, 16, 13-29.
[11] Hone, D. W. E., & Naish, D. (2013). The ‘species recognition hypothesis’ does not explain the presence and evolution of exaggerated structures in non‐avialan dinosaurs. Journal of Zoology, 290(3), 172-180.
[12] Knapp, A., Knell, R. J., Farke, A. A., Loewen, M. A., & Hone, D. W. (2018). Patterns of divergence in the morphology of ceratopsian dinosaurs: sympatry is not a driver of ornament evolution . Proceedings of the Royal Society B: Biological Sciences, 285(1875), 20180312.
[13] Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. Tyrannosaurus rex the Tyrant king, 355-370.
[14] Farke, A. A., Wolff, E. D., & Tanke, D. H. (2009). Evidence of combat in Triceratops. PLoS One, 4(1), e4252.
[15] Farlow, J. O. (1990). Dynamic Dinosaurs-Dynamics of Dinosaurs and Other Extinct Giants. R. McNeill Alexander. Columbia University Press; New York. 1989. viii+ 167 pp. $30.00. Paleobiology, 16(2), 234-241.
[16] Reid, R.E.H. (1997). "Histology of bones and teeth". In Currie, P. J.; Padian, K. (eds.). Encyclopedia of Dinosaurs. San Diego, CA.: Academic Press. pp. 329–339.
[17] Horner, J. R., & Goodwin, M. B. (2009). Extreme cranial ontogeny in the Upper Cretaceous dinosaur Pachycephalosaurus. PLoS One, 4(10), e7626.
[18] Horner, J. R., & Lamm, E. T. (2011). Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis. Comptes Rendus Palevol, 10(5-6), 439-452.
[19] Erickson, G. M., Sidebottom, M. A., Kay, D. I., Turner, K. T., Ip, N., Norell, M. A., ... & Krick, B. A. (2015). Wear biomechanics in the slicing dentition of the giant horned dinosaur Triceratops. Science Advances, 1(5), e1500055.
[20] Dodson, P.; Forster, C. A.; Sampson, S. D. (2004). "Ceratopsidae". In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (second ed.). Berkeley: University of California Press. pp. 494–513. ISBN 978-0-520-24209-8.
[21] Erickson, G. M., Krick, B. A., Hamilton, M., Bourne, G. R., Norell, M. A., Lilleodden, E., & Sawyer, W. G. (2012). Complex dental structure and wear biomechanics in hadrosaurid dinosaurs. Science, 338(6103), 98-101.
[22] markwitton-com.blogspot.com/2015/12/dinosaur-scales-some-thoughts-for.html
[23] Horner, J. R., & Goodwin, M. B. (2006). Major cranial changes during Triceratops ontogeny. Proceedings of the Royal Society B: Biological Sciences, 273(1602), 2757-2761.