Post by dinosauria101 on Apr 25, 2019 6:55:23 GMT 5
Acrocanthosaurus atokensis

Reconstruction of Acrocanthosaurus atokensis. © @ bluegekkk0->.
Temporal range: Early Cretaceous (Aptian-Albian)[1][2]; >112-105 Ma[3][4]
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Olfactores
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Class: Reptilia
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Order: Saurischia
Clade: Eusaurischia
Suborder: Theropoda
Clade: Neotheropoda
Clade: Averostra
Clade: Tetanurae
Clade: Orionides
Clade: Avetheropoda
Infraorder: †Carnosauria
Superfamily: †Allosauroidea
Clade: †Carcharodontosauria
Family: †Carcharodontosauridae
Genus: †Acrocanthosaurus
Species: †A. atokensis
Acrocanthosaurus is a genus of carcharodontosaurian theropod that lived in North America from the Aptian to Albian ages of the Early Cretaceous. It only contains one species, A. atokensis.
Habitat:
Based off of the rocks and fossils in what is now Oklahoma (formerly an isthmus), the average yearly temperature of Acrocanthosaurus’ habitat would have been 32oC (90oF), with the summer months most likely having a temperature in the high 90s to low 100s Fahrenheit (well over 30oC). Throughout the existence of the species, the precursor to the Gulf of Mexico slowly rose such that, by 110 million years ago, the coastline reached southeastern Oklahoma. At the time Acrocanthosaurus lived, days would have been slightly shorter than today (~23 hours) and years would have been slightly longer (~369 days) due to the slightly greater proximity of the Sun, Earth, and Moon.[5]
Fossil charcoal found in the same rock formations that Acrocanthosaurus fossils were found in indicate occasional wildfires throughout the habitat (possibly caused by lightning strikes). In the Twin Mountains and Glen Rose Formations, angiosperms were present, but rare, with conifers being dominant. Giant Frenelopsis trees were especially prominent, and their needle-like leaf morphology suggests that the environment was arid, not jungle-like.[5]
Description:
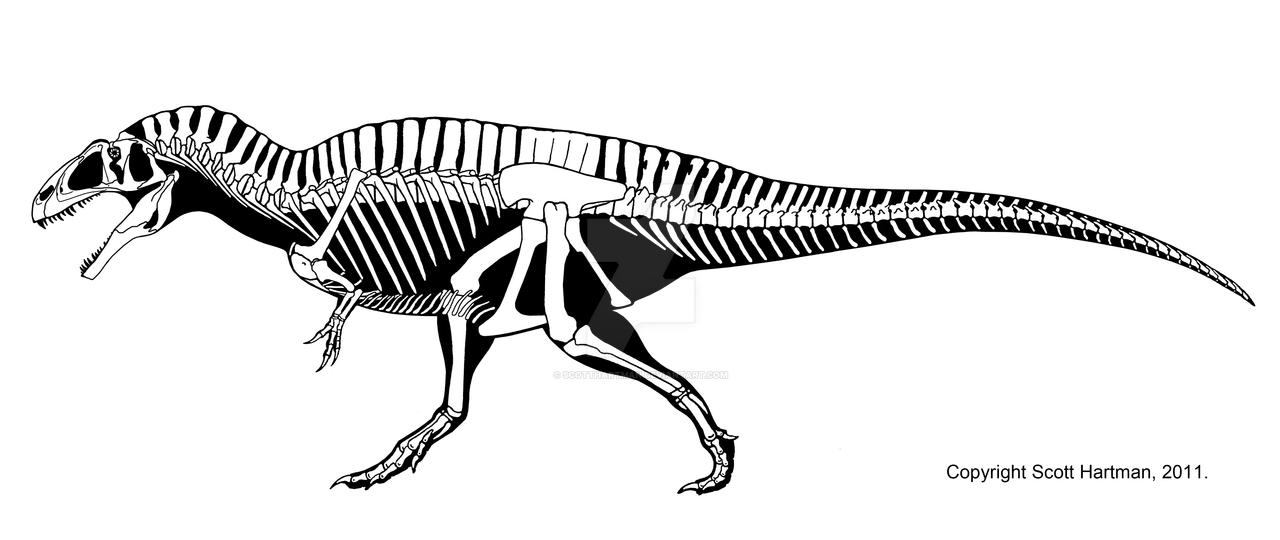
For the most part, Acrocanthosaurus possessed the typical theropod bauplan. However, its most distinctive feature (which it was named for) are the tall neural spines of its vertebrae.[6] The skull was large; specimen NCSM 14345 had a skull measuring almost 129 cm long.[7] Although the forelimbs were short, they were extremely robust, and the only theropods with stockier forelimbs were the megalosauroids.[8] The three manual unguals on each manus were raptor-like talons.[5] The lower leg bones were rather short in comparison to the length of the femur (~5.8% shorter than expected).[9] The foot is comparable to that of Allosaurus or Sinraptor, but more robust. The pedal unguals were triangular in cross section and similar to those of other carnosaurs.[7]
Gregory S. Paul estimated the animal to have been 11 meters long and around 4.4 tonnes.[10]
Biology:
Acrocanthosaurus is the only unequivocal taxon of gigantic theropod in its ecosystem. Parallels have been drawn between the end-Early and end-Late Cretaceous North American ecosystems, where there only seems to have been one extremely large-bodied predatory theropod (Acrocanthosaurus and Tyrannosaurus in the end Albian and Maastrichtian, respectively).[4] Acrocanthosaurus was possibly a keystone species[4], and an apex predator. It would have taken ~20-25 years to reach the aforementioned[10] elephantine sizes, but ~18-24 years to reach >3 tonnes.[4]
The teeth were ziphodont. The posterior edges of Acrocanthosaurus teeth have 55 serrations per inch. This is greater than the amount of serrations Tyrannosaurus had on its teeth (45 per inch). If saw blades are an appropriate functional analogy, this would indicate that Acrocanthosaurus’ teeth were not as specialized at cutting through bone as the teeth of Tyrannosaurus.[5] However, Acrocanthosaurus still possessed a powerful slicing bite. Force profiles of the mandible of Acrocanthosaurus indicate that bite force decreased slowly in the posterior half of the tooth row. The mandible was stronger in both dorsoventral and lateromedial planes at the second alveolus than at the middentary. The mandibles were extremely well buttressed dorsoventrally in the posterior half of the tooth row, but were much rounder in the anterior region, suggesting that lateromedial loads were much more important in the anterior end of the jaw than at the posterior. These features indicate that Acrocanthosaurus had a powerful slicing bite (once estimated to be 3.79 times stronger than the bite force of an American alligator; Alligator mississippiensis), and could manipulate prey with the anterior ends of their jaws.[11]

The skull of Acrocanthosaurus. © @ Ben Miller->.
While Acrocanthosaurus would have been able to manually grasp prey animals and body parts (of larger prey) below its chest, it would not have been able to grasp anything above its chest due to the forelimbs’ limited anterior range of motion[8] (although, see [12] for a study suggesting that estimates of dinosaur forelimb range of motion are too conservative; this would suggest Acrocanthosaurus could move its forelimbs somewhat farther upwards than [8] estimated). Prey items were therefore first seized orally. Digits I and II converged during flexion, and the claw on digit I (and possibly II) was permanently and strongly flexed. Only digit III was capable of abduction or adduction, but all three fingers were capable of extreme hyperextension. The muscle scars on Acrocanthosaurus’ forelimbs suggest the arms were heavily muscled and made for strenuous activity (e.g. wrestling with large, violently struggling prey). The fact that the first and possibly second claws were permanently flexed meant that a struggling prey item attempting to escape would only have further impaled itself on the claws further. After capturing prey with the jaws, Acrocanthosaurus could have either held on with its forelimbs and dispatched it with its powerful slicing bite, or held on with its jaws and delivered fatal slashing blows.[8] The shortened condition of the forelimbs were diagnosed as anterior micromelias.[13]

Right forelimb of Acrocanthosaurus. © @ Black Hills Institute of Geological Research, Inc.->.
The function of the tall neural spines appears to remain unknown. Functions include anchoring of extensive musculature (forming a muscular hump), fat storage, thermoregulation, and species recognition. At least most of these proposals have objections to them.[5]
Acrocanthosaurus lacked cranial ornamentation. Research indicates that theropod lineages that had cranial ornamentation evolved gigantic (≥1,000 kg) body size more often and an order of magnitude more rapidly than those without. Therefore, Acrocanthosaurus represents a deviation against the general trend of head-ornamented theropods more often evolving gigantism.[14]
Acrocanthosaurus’ large body size can be attributed to a habit of hunting the large sauropod dinosaurs it coexisted with. Earlier Jurassic theropods show evidence of increasing in size as the sauropods increased in size. In the same manner, the enormous size of Acrocanthosaurus (as well as the thicker, stronger neck and longer, deeper skull compared to previous allosaurs, and well-braced dorsal spine) would seem to indicate that it had evolved to tackle large sauropods. Previously, there have been suggestions that large theropod and sauropod footprints (attributable to Acrocanthosaurus and Sauroposeidon, respectively) at Glen Rose and Davenport Ranch are direct evidence for predator prey interactions. One set of tracks has been suggested to have been the result of an Acrocanthosaurus grappling with the base of its prey’s tail. However, this is now considered outdated, as it would have been impossible for the theropod to go over and around the prey’s tail while running.[15]
The relatively short lower legs of Acrocanthosaurus suggest it was not a particularly cursorial predator.[9]
Response to environmental changes:
There is a proposal that as the precursor to the Gulf of Mexico expanded and reached the Oklahoma-Texas border, the environment between the sea and the Ouachita Mountains became too constricted. This might have forced larger dinosaurs out of the area. Given the presence of Acrocanthosaurus further up north in the Cloverly Formation at a slightly younger date[4][5], this might indicate that it migrated up north.[5] Acrocanthosaurus appears to have gone extinct shortly afterwards.[5]
References:
[1] Cifelli, R. L., Gardner, J. D., Nydam, R. L., & Brinkman, D. L. (1997). Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes, 57(4), 124-131.
[2] Weishampel, D. B., Dodson, P., & Osmólska, H. (Eds.). (2007). The dinosauria. Univ of California Press.
[3] Adams, T. L. (2013). A new neosuchian crocodyliform from the Lower Cretaceous (Late Aptian) Twin Mountains Formation of north-central Texas. Journal of Vertebrate Paleontology, 33(1), 85-101. (Upper age range is based upon a time scale in this paper taken from Gradstein et al. (2004))
[4] Michael, D. D., Melstrom, K. M., & Eddy, D. R. (2012). Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis. Palaeogeography, Palaeoclimatology, Palaeoecology, 333, 13-23.
[5] Carpenter, K. (2016). Acrocanthosaurus Inside and Out. University of Oklahoma Press.
[6] Stovall, J. W., & Langston, W. (1950). Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. The American Midland Naturalist, 43(3), 696-728.
[7] Currie, P. J., & Carpenter, K. (2000). A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the lower Cretaceous Antlers formation (lower Cretaceous, Aptian) of Oklahoma, USA. Geodiversitas, 22(2), 207-246.
[8] Senter, P., & Robins, J. H. (2005). Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour. Journal of Zoology, 266(3), 307-318.
[9] Persons IV, W. S., & Currie, P. J. (2016). An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry. Scientific reports, 6(1), 1-12.
[10] Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 102
[11] Therrien, F., Henderson, D. M., & Ruff, C. B. (2005). Bite me: biomechanical models of theropod mandibles and implications for feeding behavior. The carnivorous dinosaurs, 179-237.
[12] Hutson, J. D., & Hutson, K. N. (2013). Using the American alligator and a repeated-measures design to place constraints on in vivo shoulder joint range of motion in dinosaurs and other fossil archosaurs. Journal of Experimental Biology, 216(2), 275-284.
[13] Guinard, G. (2020). Forelimb shortening of Carcharodontosauria (Dinosauria: Theropoda): an update on evolutionary anterior micromelias in non-avian theropods. Zoology, 139, 125756.
[14] Gates, T. A., Organ, C., & Zanno, L. E. (2016). Bony cranial ornamentation linked to rapid evolution of gigantic theropod dinosaurs. Nature communications, 7(1), 1-10.
[15] Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs: life in the age of giants. JHU Press.
Reconstruction of Acrocanthosaurus atokensis. © @ bluegekkk0->.
Temporal range: Early Cretaceous (Aptian-Albian)[1][2]; >112-105 Ma[3][4]
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Olfactores
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Class: Reptilia
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Order: Saurischia
Clade: Eusaurischia
Suborder: Theropoda
Clade: Neotheropoda
Clade: Averostra
Clade: Tetanurae
Clade: Orionides
Clade: Avetheropoda
Infraorder: †Carnosauria
Superfamily: †Allosauroidea
Clade: †Carcharodontosauria
Family: †Carcharodontosauridae
Genus: †Acrocanthosaurus
Species: †A. atokensis
Acrocanthosaurus is a genus of carcharodontosaurian theropod that lived in North America from the Aptian to Albian ages of the Early Cretaceous. It only contains one species, A. atokensis.
Habitat:
Based off of the rocks and fossils in what is now Oklahoma (formerly an isthmus), the average yearly temperature of Acrocanthosaurus’ habitat would have been 32oC (90oF), with the summer months most likely having a temperature in the high 90s to low 100s Fahrenheit (well over 30oC). Throughout the existence of the species, the precursor to the Gulf of Mexico slowly rose such that, by 110 million years ago, the coastline reached southeastern Oklahoma. At the time Acrocanthosaurus lived, days would have been slightly shorter than today (~23 hours) and years would have been slightly longer (~369 days) due to the slightly greater proximity of the Sun, Earth, and Moon.[5]
Fossil charcoal found in the same rock formations that Acrocanthosaurus fossils were found in indicate occasional wildfires throughout the habitat (possibly caused by lightning strikes). In the Twin Mountains and Glen Rose Formations, angiosperms were present, but rare, with conifers being dominant. Giant Frenelopsis trees were especially prominent, and their needle-like leaf morphology suggests that the environment was arid, not jungle-like.[5]
Description:
For the most part, Acrocanthosaurus possessed the typical theropod bauplan. However, its most distinctive feature (which it was named for) are the tall neural spines of its vertebrae.[6] The skull was large; specimen NCSM 14345 had a skull measuring almost 129 cm long.[7] Although the forelimbs were short, they were extremely robust, and the only theropods with stockier forelimbs were the megalosauroids.[8] The three manual unguals on each manus were raptor-like talons.[5] The lower leg bones were rather short in comparison to the length of the femur (~5.8% shorter than expected).[9] The foot is comparable to that of Allosaurus or Sinraptor, but more robust. The pedal unguals were triangular in cross section and similar to those of other carnosaurs.[7]
Gregory S. Paul estimated the animal to have been 11 meters long and around 4.4 tonnes.[10]
Biology:
Acrocanthosaurus is the only unequivocal taxon of gigantic theropod in its ecosystem. Parallels have been drawn between the end-Early and end-Late Cretaceous North American ecosystems, where there only seems to have been one extremely large-bodied predatory theropod (Acrocanthosaurus and Tyrannosaurus in the end Albian and Maastrichtian, respectively).[4] Acrocanthosaurus was possibly a keystone species[4], and an apex predator. It would have taken ~20-25 years to reach the aforementioned[10] elephantine sizes, but ~18-24 years to reach >3 tonnes.[4]
The teeth were ziphodont. The posterior edges of Acrocanthosaurus teeth have 55 serrations per inch. This is greater than the amount of serrations Tyrannosaurus had on its teeth (45 per inch). If saw blades are an appropriate functional analogy, this would indicate that Acrocanthosaurus’ teeth were not as specialized at cutting through bone as the teeth of Tyrannosaurus.[5] However, Acrocanthosaurus still possessed a powerful slicing bite. Force profiles of the mandible of Acrocanthosaurus indicate that bite force decreased slowly in the posterior half of the tooth row. The mandible was stronger in both dorsoventral and lateromedial planes at the second alveolus than at the middentary. The mandibles were extremely well buttressed dorsoventrally in the posterior half of the tooth row, but were much rounder in the anterior region, suggesting that lateromedial loads were much more important in the anterior end of the jaw than at the posterior. These features indicate that Acrocanthosaurus had a powerful slicing bite (once estimated to be 3.79 times stronger than the bite force of an American alligator; Alligator mississippiensis), and could manipulate prey with the anterior ends of their jaws.[11]
The skull of Acrocanthosaurus. © @ Ben Miller->.
While Acrocanthosaurus would have been able to manually grasp prey animals and body parts (of larger prey) below its chest, it would not have been able to grasp anything above its chest due to the forelimbs’ limited anterior range of motion[8] (although, see [12] for a study suggesting that estimates of dinosaur forelimb range of motion are too conservative; this would suggest Acrocanthosaurus could move its forelimbs somewhat farther upwards than [8] estimated). Prey items were therefore first seized orally. Digits I and II converged during flexion, and the claw on digit I (and possibly II) was permanently and strongly flexed. Only digit III was capable of abduction or adduction, but all three fingers were capable of extreme hyperextension. The muscle scars on Acrocanthosaurus’ forelimbs suggest the arms were heavily muscled and made for strenuous activity (e.g. wrestling with large, violently struggling prey). The fact that the first and possibly second claws were permanently flexed meant that a struggling prey item attempting to escape would only have further impaled itself on the claws further. After capturing prey with the jaws, Acrocanthosaurus could have either held on with its forelimbs and dispatched it with its powerful slicing bite, or held on with its jaws and delivered fatal slashing blows.[8] The shortened condition of the forelimbs were diagnosed as anterior micromelias.[13]
Right forelimb of Acrocanthosaurus. © @ Black Hills Institute of Geological Research, Inc.->.
The function of the tall neural spines appears to remain unknown. Functions include anchoring of extensive musculature (forming a muscular hump), fat storage, thermoregulation, and species recognition. At least most of these proposals have objections to them.[5]
Acrocanthosaurus lacked cranial ornamentation. Research indicates that theropod lineages that had cranial ornamentation evolved gigantic (≥1,000 kg) body size more often and an order of magnitude more rapidly than those without. Therefore, Acrocanthosaurus represents a deviation against the general trend of head-ornamented theropods more often evolving gigantism.[14]
Acrocanthosaurus’ large body size can be attributed to a habit of hunting the large sauropod dinosaurs it coexisted with. Earlier Jurassic theropods show evidence of increasing in size as the sauropods increased in size. In the same manner, the enormous size of Acrocanthosaurus (as well as the thicker, stronger neck and longer, deeper skull compared to previous allosaurs, and well-braced dorsal spine) would seem to indicate that it had evolved to tackle large sauropods. Previously, there have been suggestions that large theropod and sauropod footprints (attributable to Acrocanthosaurus and Sauroposeidon, respectively) at Glen Rose and Davenport Ranch are direct evidence for predator prey interactions. One set of tracks has been suggested to have been the result of an Acrocanthosaurus grappling with the base of its prey’s tail. However, this is now considered outdated, as it would have been impossible for the theropod to go over and around the prey’s tail while running.[15]
The relatively short lower legs of Acrocanthosaurus suggest it was not a particularly cursorial predator.[9]
Response to environmental changes:
There is a proposal that as the precursor to the Gulf of Mexico expanded and reached the Oklahoma-Texas border, the environment between the sea and the Ouachita Mountains became too constricted. This might have forced larger dinosaurs out of the area. Given the presence of Acrocanthosaurus further up north in the Cloverly Formation at a slightly younger date[4][5], this might indicate that it migrated up north.[5] Acrocanthosaurus appears to have gone extinct shortly afterwards.[5]
References:
[1] Cifelli, R. L., Gardner, J. D., Nydam, R. L., & Brinkman, D. L. (1997). Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes, 57(4), 124-131.
[2] Weishampel, D. B., Dodson, P., & Osmólska, H. (Eds.). (2007). The dinosauria. Univ of California Press.
[3] Adams, T. L. (2013). A new neosuchian crocodyliform from the Lower Cretaceous (Late Aptian) Twin Mountains Formation of north-central Texas. Journal of Vertebrate Paleontology, 33(1), 85-101. (Upper age range is based upon a time scale in this paper taken from Gradstein et al. (2004))
[4] Michael, D. D., Melstrom, K. M., & Eddy, D. R. (2012). Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis. Palaeogeography, Palaeoclimatology, Palaeoecology, 333, 13-23.
[5] Carpenter, K. (2016). Acrocanthosaurus Inside and Out. University of Oklahoma Press.
[6] Stovall, J. W., & Langston, W. (1950). Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. The American Midland Naturalist, 43(3), 696-728.
[7] Currie, P. J., & Carpenter, K. (2000). A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the lower Cretaceous Antlers formation (lower Cretaceous, Aptian) of Oklahoma, USA. Geodiversitas, 22(2), 207-246.
[8] Senter, P., & Robins, J. H. (2005). Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour. Journal of Zoology, 266(3), 307-318.
[9] Persons IV, W. S., & Currie, P. J. (2016). An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry. Scientific reports, 6(1), 1-12.
[10] Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 102
[11] Therrien, F., Henderson, D. M., & Ruff, C. B. (2005). Bite me: biomechanical models of theropod mandibles and implications for feeding behavior. The carnivorous dinosaurs, 179-237.
[12] Hutson, J. D., & Hutson, K. N. (2013). Using the American alligator and a repeated-measures design to place constraints on in vivo shoulder joint range of motion in dinosaurs and other fossil archosaurs. Journal of Experimental Biology, 216(2), 275-284.
[13] Guinard, G. (2020). Forelimb shortening of Carcharodontosauria (Dinosauria: Theropoda): an update on evolutionary anterior micromelias in non-avian theropods. Zoology, 139, 125756.
[14] Gates, T. A., Organ, C., & Zanno, L. E. (2016). Bony cranial ornamentation linked to rapid evolution of gigantic theropod dinosaurs. Nature communications, 7(1), 1-10.
[15] Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs: life in the age of giants. JHU Press.