


Post by Runic on Aug 27, 2013 22:28:49 GMT 5
I've been researching for a while now and have composed a thread displaying the killing tactic and logic behind the evolved killing weapons of "Modern" birds of prey. Either read it all (Strongly suggest) or if your lazy scroll to the bottom for a summary. First study was done on kestrel It's mainly divided into 3 parts.
Usage of the feet and talons for killing between large and small prey specialists, difference in foot morphology and grip strength, and consumption method.
---------------------------------------------------------------------------------------------------------------------------
www.jstor.org/discover/pgs/index?id=10.2307/3677335&img=dtc.12.tif.gif&uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097&orig=/discover/10.2307/3677335?uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097
The technique used by birds of prey to kill their quarry has rarely been described in detail. Although it is common to watch a raptor hunting, distance and natural obstacles normally prevent the observer from observing what actually happens to the prey after being grasped by the bird. Therefore, we studied the predatory sequence and killing technique displayed by a sample of ten wild Eurasian Kestrels temporarily held in captivity because of various injuries. They were offered individually either a laboratory mouse or young laboratory rat. Direct observation was complemented by necropsy of the prey confirming that bill strikes (Biting with the beak) frequently observed after grasping the prey are not used to kill it but merely to damage its central nervous system thereby minimizing escape attempts. Furthermore, talons are not used as weapons at all, but only as tools that elongate the toes to hold the quarry more firmly. In contrast, several signs indicating death by suffocation were regularly found and these were confirmed by inspection of rodents sacrificed by CO2. We also discuss a hypothesis explaining the adaptive and evolutionary significance of the kestrel's use of toe pressure to kill the prey rather than using its obvious weapons.
www.jstor.org/discover/pgs/index?id=10.2307/1369762&img=dtc.171.tif.gif&uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097&orig=/discover/10.2307/1369762?uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097
We quantified foot squeezing pressure exerted by Common Buzzards (Buteo buteo) and Eurasian Kestrels (Falco tinnunculus) while grasping a rodent. The birds were offered either a dead laboratory mouse, a fake mouse consisting of a laboratory mouse skin surrounding a rubber pipe connected to a pressure transducer, or a live laboratory mouse. Direct observations and necropsy of the depredated mice confirm death by suffocation from the raptor's grasp. The two raptor species differed in technique of constricting the fake mouse: the buzzard relied on strong, but very brief squeezing bouts, whereas the kestrel compensated for less squeezing strength by performing constriction with prolonged duration. When observed, bites to the head were consistent with immobilizing live prey but not killing it. In contrast, squeezing by thoracic compression is sufficient to kill the prey without the need to use beak or talons.
More of this can be seen from this chart and quote,
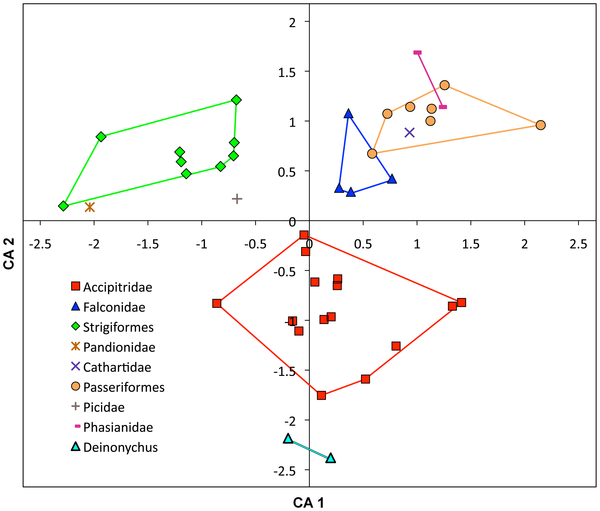
What Does This Mean?
- The small prey specialists evolved smaller talons and shorter flexor tendon to aid in the capture and constriction of the small prey the hunt.
- The large mammal and fish specialist birds of prey evolved more robust feet with relatively short and robust tarsi (foot bones towards the toes) with longer and thicker talons.
- The medium sized mammal specialist have a somewhat median of both, evolving long digits and a large digit 1 talon to equate for faster movement and larger talons for pinning etc of larger prey.
- Accipitridae (hawk and eagles mainly) have longer claws than all other family of birds but also have the weakest grip strength but in turn have wider and quicker foot movement. The reverse can be said for falconidae (falcons) and strigiformes (owls)
- Birds with smaller claws generally have a stronger pressure out-put than animals with larger and longer claws. Owls are a good example of this as they use their feet to suffocate the small prey that they capture.
-----------------------------------------------------------------------------------------------------------------
Since owls and falcons rely more on the strength of their grip to kill prey via suffocation, it would seem that birds from the family Accipitidae would rely more on the mechanical damage from their claws to kill animals their size or larger (they kill animals of sub-equal size through the RPR method. A good example of the reliance of mechanical damage from claws to take down large game would be the golden eagle predation on deer.
Quote from 221extra








^ A prime example of the mechanical damage done by the large claws of a bird through stabbing.



What can be made from this?
-Smaller prey specialists like owls and falcons rely on their grip strength for killing rather than their claws (the claws are used to keep the prey from running) while larger prey specialists like eagles and hawks mainly rely on the mechanical damage done by their claws (through stabbing) rather than the pressure exerted from their feet. For smaller prey since Accipitridae lacks the pressure sufficient enough to constrict, they simply use their large claws to hold down smaller prey while they eat it alive.
- Owls and falcons use their beaks to bite down rather than nip on their prey to immobilize the nervous system and prevent escape rather than for killing.
- Due to their foot morphology owls and falcon are incapable of taking large game on the same level as eagle and hawks.
-Due to their foot morphology eagles and hawks are able to tackle much larger prey to compensate for their lacking in the ability of catching smaller fleet footed prey that animals like owls frequently take.
-----------------------------------------------------------------------------------------------------------------------------
SUMMARY
- Accipitridae have longer claws than other birds of prey but in return have weaker grip strengths thus rely more on mechanical damage done by their claws via stabbing (On larger prey) and biting and eating alive (RPR method)
rather than suffocation of prey.
-Smaller prey specialists like owls and falcons rely on their grip strength for killing rather than their claws (the claws are used to keep the prey from running) while larger prey specialists like eagles and hawks mainly rely on the mechanical damage done by their claws (through stabbing) rather than the pressure exerted from their feet. For smaller prey since Accipitridae lacks the pressure sufficient enough to constrict, they simply use their large claws to hold down smaller prey while they eat it alive.
- Owls and falcon have shorter talons but stronger grip strength than eagle and hawks
- Falcon and owl rely more on constriction of prey and nervous system immobilation to kill why acccipitridae rely on mechanical damage via stabbing of large prey or using their large claws for pinning of small prey for live consumption.
Larger prey specialist have larger talons and longer flexor tendon while small prey specialist have it reversed.
- Accipitridae kill larger prey via stabbing of the backbone and head.
- Birds specialized in catching other birds have a medium between grip strength and talon length.
For more info on bird bite force, see here
onlinelibrary.wiley.com/doi/10.1111/j.1365-2435.2008.01494.x/full
Usage of the feet and talons for killing between large and small prey specialists, difference in foot morphology and grip strength, and consumption method.
---------------------------------------------------------------------------------------------------------------------------
www.jstor.org/discover/pgs/index?id=10.2307/3677335&img=dtc.12.tif.gif&uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097&orig=/discover/10.2307/3677335?uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097
The technique used by birds of prey to kill their quarry has rarely been described in detail. Although it is common to watch a raptor hunting, distance and natural obstacles normally prevent the observer from observing what actually happens to the prey after being grasped by the bird. Therefore, we studied the predatory sequence and killing technique displayed by a sample of ten wild Eurasian Kestrels temporarily held in captivity because of various injuries. They were offered individually either a laboratory mouse or young laboratory rat. Direct observation was complemented by necropsy of the prey confirming that bill strikes (Biting with the beak) frequently observed after grasping the prey are not used to kill it but merely to damage its central nervous system thereby minimizing escape attempts. Furthermore, talons are not used as weapons at all, but only as tools that elongate the toes to hold the quarry more firmly. In contrast, several signs indicating death by suffocation were regularly found and these were confirmed by inspection of rodents sacrificed by CO2. We also discuss a hypothesis explaining the adaptive and evolutionary significance of the kestrel's use of toe pressure to kill the prey rather than using its obvious weapons.
www.jstor.org/discover/pgs/index?id=10.2307/1369762&img=dtc.171.tif.gif&uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097&orig=/discover/10.2307/1369762?uid=3739616&uid=2&uid=4&uid=3739256&sid=21102175500097
We quantified foot squeezing pressure exerted by Common Buzzards (Buteo buteo) and Eurasian Kestrels (Falco tinnunculus) while grasping a rodent. The birds were offered either a dead laboratory mouse, a fake mouse consisting of a laboratory mouse skin surrounding a rubber pipe connected to a pressure transducer, or a live laboratory mouse. Direct observations and necropsy of the depredated mice confirm death by suffocation from the raptor's grasp. The two raptor species differed in technique of constricting the fake mouse: the buzzard relied on strong, but very brief squeezing bouts, whereas the kestrel compensated for less squeezing strength by performing constriction with prolonged duration. When observed, bites to the head were consistent with immobilizing live prey but not killing it. In contrast, squeezing by thoracic compression is sufficient to kill the prey without the need to use beak or talons.
Each bird wa s tested onl y once , in orde r to avoid an y modification s in behaviour du e to repetition o f the same test . A tota l o f 27 Buzzards were used. Among these , 10 were adults and 17 juveniles. The tests were carried out i n the morning , between March an d July . An adul t laboratory mouse (Mus domesticus) of Agouti phenotype was offered as prey. This mouse was the firs t they coul d approac h afte r arriva l a t th e CRR . Th e mous e wa s inserted manually into a hole under the platform. Fifteen minutes later it was elevated semi-automatically onto the platform by the observer (thi s marked the start of the observation period) but only if the Buzzard was on the perch. The mouse was unable to escape and was completely visible to the Buzzard. We kept the number o f tests as low as possible compatible with procuring a significant sample , whils t a t th e sam e tim e sacrificin g a s fe w mic e a s necessary, as recommended fo r instance by Huntingford (1984).
The birds were offered the test-prey after four days of fasting. We wished to enhanc e an d equalis e th e predator y motivatio n fo r al l birds . I n fact , preliminary tests carried ou t afte r only two days without food rarely caused prédation. On the other hand it has already been demonstrated that hunger is correlated wit h pre y killing i n captiv e raptors , suc h a s the Broad-winged
135
Hawk (Buteo platypterus), the American Kestre l (Falco sparverius), and Screech-owl (Otus asioj Muelle r 1973; Marti & Hogue 1979).
The ag e o f the birds , i.e . whethe r adul t (AD ) o r juvenil e (JU ) wa s considered, but the sex was not taken into account , due to the difficulty of detecting thi s fo r certai n i n subadults . Al l th e bird s were experiencin g captivity fo r the first time.
The observation sessio n wa s stopped i f no prédation occurred afte r 60 minutes from the appearance o f the mouse on the platform . The behaviour patterns displayed by the birds before and during prédation were recorded.
RESULTS
Only 17 birds took the prey (5 AD and 12 JU) (Chi-square test, p > 0.1 ) (Table 1). The behaviour before prédation can be classified as either active or non-active. Eleven birds , ou t o f the 17 predating, remained totally inactive on th e perch , showin g no differenc e between ag e classes , whil e only two remained inactiv e amon g thos e tha t di d no t tak e th e mouse . Preening , performed by eight individuals, was mostly followed by feather shaking, with a significant correlation between the two patterns (C contingency coefficient = 0.524 , p < 0.001 ) (Siege l 1956) . Nevertheless, preening wa s performed irrespective o f age.
Preening wa s performed with a mean latency o f 1296.0 + 278. 5 (SEM) seconds, bu t wit h n o differenc e between JU an d AD birds . Moreover , preening was negatively correlated (C = 0.375 , p < 0.05 ) with subsequent prédation. The only three Buzzards performing both preening and prédation clearly showed a direc t trend fo r the latencies , i.e . the earlie r th e preening time, the earlier the prédation.
Some individuals were also observed to abandon perch contact with one leg, pushing this forward and closing the talons, as when taking something with the foot . On othe r occasion s some birds performed fo r a certain time rhythmic vertical movements with the head. Unfortunately it was impossible to quantify the frequency and duration o f these patterns.
Among the birds active during the tes t 12 individuals performed one or more flights within the pen. These were not correlated either with age or with eventual capture o f the prey.
136
Table 1: The total number o f birds tested an d the proportion o f those taking or not taking the mouse . For both groups the number o f birds displaying o r not the various behaviour patterns considered.
No. o f birds tested 27
No. o f birds preying 17
No. o f birds not preying 10
No. o f birds preying and: - preening/feather shaking 2 - not preening/feather shaking 15 - moving on the perch 4 - not moving on the perch 13 - flying 7 - not flying 10 - totally inactive before prédation 11
No. o f birds not preying and: - preening/feather shaking 6 - not preening/feather shaking 4 - moving on the perch 5 - not moving on the perch 5 - flying 5 - not flying 5 - totally inactive during the test 2
The mouse was approached in two different ways: 1) directly , i.e. landing directly on the prey (Fig . 2); 2) indirectly, i.e. landing on the platform and then walking toward the mouse and capturing it with an extended foot . The direct approach, most often used, was recorded in 13 birds (8 JU and 5 AD), while the indirect one was performed more rarely (4 cases) and only by JU individuals.
137
The birds were offered the test-prey after four days of fasting. We wished to enhanc e an d equalis e th e predator y motivatio n fo r al l birds . I n fact , preliminary tests carried ou t afte r only two days without food rarely caused prédation. On the other hand it has already been demonstrated that hunger is correlated wit h pre y killing i n captiv e raptors , suc h a s the Broad-winged
135
Hawk (Buteo platypterus), the American Kestre l (Falco sparverius), and Screech-owl (Otus asioj Muelle r 1973; Marti & Hogue 1979).
The ag e o f the birds , i.e . whethe r adul t (AD ) o r juvenil e (JU ) wa s considered, but the sex was not taken into account , due to the difficulty of detecting thi s fo r certai n i n subadults . Al l th e bird s were experiencin g captivity fo r the first time.
The observation sessio n wa s stopped i f no prédation occurred afte r 60 minutes from the appearance o f the mouse on the platform . The behaviour patterns displayed by the birds before and during prédation were recorded.
RESULTS
Only 17 birds took the prey (5 AD and 12 JU) (Chi-square test, p > 0.1 ) (Table 1). The behaviour before prédation can be classified as either active or non-active. Eleven birds , ou t o f the 17 predating, remained totally inactive on th e perch , showin g no differenc e between ag e classes , whil e only two remained inactiv e amon g thos e tha t di d no t tak e th e mouse . Preening , performed by eight individuals, was mostly followed by feather shaking, with a significant correlation between the two patterns (C contingency coefficient = 0.524 , p < 0.001 ) (Siege l 1956) . Nevertheless, preening wa s performed irrespective o f age.
Preening wa s performed with a mean latency o f 1296.0 + 278. 5 (SEM) seconds, bu t wit h n o differenc e between JU an d AD birds . Moreover , preening was negatively correlated (C = 0.375 , p < 0.05 ) with subsequent prédation. The only three Buzzards performing both preening and prédation clearly showed a direc t trend fo r the latencies , i.e . the earlie r th e preening time, the earlier the prédation.
Some individuals were also observed to abandon perch contact with one leg, pushing this forward and closing the talons, as when taking something with the foot . On othe r occasion s some birds performed fo r a certain time rhythmic vertical movements with the head. Unfortunately it was impossible to quantify the frequency and duration o f these patterns.
Among the birds active during the tes t 12 individuals performed one or more flights within the pen. These were not correlated either with age or with eventual capture o f the prey.
136
Table 1: The total number o f birds tested an d the proportion o f those taking or not taking the mouse . For both groups the number o f birds displaying o r not the various behaviour patterns considered.
No. o f birds tested 27
No. o f birds preying 17
No. o f birds not preying 10
No. o f birds preying and: - preening/feather shaking 2 - not preening/feather shaking 15 - moving on the perch 4 - not moving on the perch 13 - flying 7 - not flying 10 - totally inactive before prédation 11
No. o f birds not preying and: - preening/feather shaking 6 - not preening/feather shaking 4 - moving on the perch 5 - not moving on the perch 5 - flying 5 - not flying 5 - totally inactive during the test 2
The mouse was approached in two different ways: 1) directly , i.e. landing directly on the prey (Fig . 2); 2) indirectly, i.e. landing on the platform and then walking toward the mouse and capturing it with an extended foot . The direct approach, most often used, was recorded in 13 birds (8 JU and 5 AD), while the indirect one was performed more rarely (4 cases) and only by JU individuals.
137
It is most likely that death was caused by suffocation or by crushing, since the us e o f talons t o pierce the mouse' s body was never clearly observed . Nevertheless, pecking was most often recorded . This activity was even performed repeatedly and with a mean duration o f 6.1 + 0. 6 seconds. The pecks were not given with the tip of the bill , but by slightly opening the mandibles and giving a sort of bite. It was impossible to measure the pecking strength, but it was judged seemingly higher by us when the cephalic region was targeted, which occurred most o f the time. The flanks were more rarely "bitten" (4 times) and the hind quarters only once.
138
Prey ingestio n wa s carrie d ou t wit h th e bird s constantl y displayin g a particular "ingestion" posture (Fig. 3), different from the "blocking" posture described above , and generally a t the place of capture. Independently from the time taken for ingestion the Buzzards never lost contact with the mouse. The raptor stood over its prey , holding it with its head facing towards the Buzzard itself and both fee t "blocking" the mouse at the centre of its body. The eventual ingestion started from the mouse's head. The feet held the prey on the ground while the bill tore off scraps of flesh. The hind part was ingested whole , and no individual left any part of the mouse uneaten
138
Prey ingestio n wa s carrie d ou t wit h th e bird s constantl y displayin g a particular "ingestion" posture (Fig. 3), different from the "blocking" posture described above , and generally a t the place of capture. Independently from the time taken for ingestion the Buzzards never lost contact with the mouse. The raptor stood over its prey , holding it with its head facing towards the Buzzard itself and both fee t "blocking" the mouse at the centre of its body. The eventual ingestion started from the mouse's head. The feet held the prey on the ground while the bill tore off scraps of flesh. The hind part was ingested whole , and no individual left any part of the mouse uneaten
We quantified the extent of adaptive radiation in the evolution of the hindlimb in the bird-of-prey community on Tasmania. Assessments of the ecological capabilities of raptor species are often based on a visual inspection of their hindlimb structure, with little recourse to direct biomechanical or functional evidence. We examined the links between hindlimb structure and patterns of diet, foraging, and habitat use in raptors by analytically investigating tarsus, toe, and talon measurements. We identified groupings on the basis of dietary preference, hunting-killing technique, and phylogeny. We found that the bird-catching specialists, which possessed relatively long digits with short talons, were consistently separable. The large-mammal and fish specialists were equipped with relatively short and robust tarsi, and short digits with long and robust talons. The hindlimbs of medium-mammal specialists were characterized by long digits and a large digit 1 talon. However, the generalist group did not possess any specializations, because their structural elements were comparable to those of other dietary groups. An association was found between the ratio of toe to talon length and the selective pressures of prey capture and ease of killing. Morphological variation in this feature was consistent with phylogeny, because the Accipitridae were characterized by a larger ratio of toe to talon length on digits 1 and 2 than the Falconidae, and the diurnal raptors possessed an interdigital pattern of larger variation in ratio of toe to talon length than the nocturnal raptors. No link was found between dietary habit and tarsus length or robustness, because these features were apparently attributed to variations in hunting style. Our analysis highlights the interrelationship between the morphology of hindlimb structure and the functional pressures associated with predatory lifestyles.
Aspects de la morphologie des membres postérieurs de quelques oiseaux de proie australiens : Une étude comparative et quantitative
Aspects de la morphologie des membres postérieurs de quelques oiseaux de proie australiens : Une étude comparative et quantitative
More of this can be seen from this chart and quote,
Relative proportions of the feet vary depending on a predominantly cursorial or grasping function (Figure 5). Extant cursorial birds (e.g. ratites; Emu, Dromaius novaehollandiae, MOR OST-1299) and similar cursorially adapted dinosaur taxa (e.g. Ornithomimidae [42]; Avimimus [43]) typically exhibit a robust D-III, foreshortened distal phalanges with shorter, subequally sized lateral digits (D-II and IV) [11], [13]–[15], [17], [45]–[48]. An opposite trend is seen in the pedal digits of Deinonychosauria and basal Avialae (e.g. Archaeopteryx): D-IV becomes especially elongated, distal non-ungual phalanges are more elongated than in specialist cursors, and D-II becomes hyperextensible. these characters are consistent with grasping rather than a cursorial function [15].
Elongation of the metatarsus increases length of the flexor tendons, thus reducing mechanical advantage (and hence, grip strength), but also increases stride length (with other cursorial benefits [46], [48]) and ‘quickness’ of the feet. Extant accipitrids have an elongate metatarsus compared to other birds of prey, granting them better ability to snatch prey, an important aspect of their predatory strategy [19], [20]. Conversely, acting as the out-lever of the hindlimb, the short robust metatarsus of owls effectively reduces the length of the flexor tendons, increasing mechanical advantage; hence owls have greater force production (grip strength) than that produced by the more elongate metatarsus of accipitrids, but at a cost of less rapid movement [19]. A relatively elongate metatarsus (sometimes subarctometatarsalian) is present in basal paravians (e.g. basal troodontid Sinovenator [49]; basal dromaeosaurid Sinornithosaurus [50]; [51], [52]), indicating a cursorial habit is basal for the clade. This is further developed into a fully arctometatarsalian metatarsus in derived troodontids [5], [37], [38], [46], [53], [54]. By contrast, derived dromaeosaurids like Deinonychus [1], Saurornitholestes (MOR 660), and Velociraptor [33] lost the elongate metatarsus, instead evolving extremely short and robust metatarsi (among the shortest relative to tibia length of all non-avian theropods [55]). This suggests that the plesiomorphic cursorial metatarsus became further adapted towards cursoriality in Troodontidae whereas Dromaeosauridae reversed selection direction, specializing towards grasping strength at the expense of speed (Figure 2).
Elongation of the metatarsus increases length of the flexor tendons, thus reducing mechanical advantage (and hence, grip strength), but also increases stride length (with other cursorial benefits [46], [48]) and ‘quickness’ of the feet. Extant accipitrids have an elongate metatarsus compared to other birds of prey, granting them better ability to snatch prey, an important aspect of their predatory strategy [19], [20]. Conversely, acting as the out-lever of the hindlimb, the short robust metatarsus of owls effectively reduces the length of the flexor tendons, increasing mechanical advantage; hence owls have greater force production (grip strength) than that produced by the more elongate metatarsus of accipitrids, but at a cost of less rapid movement [19]. A relatively elongate metatarsus (sometimes subarctometatarsalian) is present in basal paravians (e.g. basal troodontid Sinovenator [49]; basal dromaeosaurid Sinornithosaurus [50]; [51], [52]), indicating a cursorial habit is basal for the clade. This is further developed into a fully arctometatarsalian metatarsus in derived troodontids [5], [37], [38], [46], [53], [54]. By contrast, derived dromaeosaurids like Deinonychus [1], Saurornitholestes (MOR 660), and Velociraptor [33] lost the elongate metatarsus, instead evolving extremely short and robust metatarsi (among the shortest relative to tibia length of all non-avian theropods [55]). This suggests that the plesiomorphic cursorial metatarsus became further adapted towards cursoriality in Troodontidae whereas Dromaeosauridae reversed selection direction, specializing towards grasping strength at the expense of speed (Figure 2).
The questions asked were (1) whether claws and toes of birds of prey are actually different from those of other perching birds, and, if so, (2) what parameter can describe such a difference. The structure of toes and claws of the first and third toe was then evaluated in three groups of birds: Falconiformes, Strigiformes and non?raptorial species. One adult male per species was considered, and, among non?raptorial birds, only species belonging to typically or partially perching families. Only one specimen per species was chosen, from museum skin bird collections. All species examined occur in the western Palearctic, according to Cramp & Simmons (1977–1994
). Discriminant Function Analysis showed clear separation of the groups. In particular, the first canonical function segregated Falconiformes from Strigiformes, whereas the second separated Strigiformes from non?raptorials. However, Falconiformes and non?raptorials partially overlapped and were not separated. The characteristics segregating Falconiformes from Strigiformes mainly concerned claw curvature, claw length in relation to toe length, and the last phalanx shape, thin or rounded. Characters contributing to segregate Strigiformes from non?raptorials were claw curvature in relation to their radius and the shape, thin or rounded, of both claws and last phalanxes. Results indicate that Strigiformes toes and claws only superficially resemble those of Falconiformes and the shape of claws and toes of Falconiformes are much more similar to those of non?raptorial species than they are to those of Strigiformes.
). Discriminant Function Analysis showed clear separation of the groups. In particular, the first canonical function segregated Falconiformes from Strigiformes, whereas the second separated Strigiformes from non?raptorials. However, Falconiformes and non?raptorials partially overlapped and were not separated. The characteristics segregating Falconiformes from Strigiformes mainly concerned claw curvature, claw length in relation to toe length, and the last phalanx shape, thin or rounded. Characters contributing to segregate Strigiformes from non?raptorials were claw curvature in relation to their radius and the shape, thin or rounded, of both claws and last phalanxes. Results indicate that Strigiformes toes and claws only superficially resemble those of Falconiformes and the shape of claws and toes of Falconiformes are much more similar to those of non?raptorial species than they are to those of Strigiformes.
Prey capture in owls and hawks is largely dependent on the biomechanics of the hindlimbs, and both limb size and grip forces potentially determine the size of prey that can be captured and the extent of possible resource partitioning among sympatric species. Morphological study of six species of sympatric raptors—the owls Otus asio, Strix varia, and Bubo virginianus; and the hawks commonly considered their diurnal “ecological equivalents,” Falco sparverius, Buteo lineatus, and Buteo jamaicensis—revealed that, in both groups, talon closure is effected by two discrete mechanisms that function together in a potentially additive or alternative fashion. Grip force measurements obtained from live owls and hawks using “hydraulic” perches showed that grip force increases exponentially with body size and that owls produce greater forces than hawks. That finding is consistent with the distinctive osteology and myology of their hindlimbs and with their hunting behavior. These data provide some understanding of the different demands of diurnal and nocturnal hunting as well as the mechanism of coexistence for those six species in eastern woodlands
What Does This Mean?
- The small prey specialists evolved smaller talons and shorter flexor tendon to aid in the capture and constriction of the small prey the hunt.
- The large mammal and fish specialist birds of prey evolved more robust feet with relatively short and robust tarsi (foot bones towards the toes) with longer and thicker talons.
- The medium sized mammal specialist have a somewhat median of both, evolving long digits and a large digit 1 talon to equate for faster movement and larger talons for pinning etc of larger prey.
- Accipitridae (hawk and eagles mainly) have longer claws than all other family of birds but also have the weakest grip strength but in turn have wider and quicker foot movement. The reverse can be said for falconidae (falcons) and strigiformes (owls)
- Birds with smaller claws generally have a stronger pressure out-put than animals with larger and longer claws. Owls are a good example of this as they use their feet to suffocate the small prey that they capture.
-----------------------------------------------------------------------------------------------------------------
Since owls and falcons rely more on the strength of their grip to kill prey via suffocation, it would seem that birds from the family Accipitidae would rely more on the mechanical damage from their claws to kill animals their size or larger (they kill animals of sub-equal size through the RPR method. A good example of the reliance of mechanical damage from claws to take down large game would be the golden eagle predation on deer.
Quote from 221extra
Golden Eagle Attacks Pronghorn Antelope in Wyoming
WILDLIFE MANAGEMENT PRO | WILDLIFE AND ENVIRONMENTAL INFO
POSTED UNDER: OUTDOOR NEWS | JANUARY 30
The golden eagle is found across North America and is the largest bird of prey found in the United States. Golden eagles occur in the greatest numbers from Alaska southward throughout the mountain and prairie habitat of the West and into Mexico. They occur in lower numbers to the east across Canada, the Great Lakes states, and the Appalachian Mountains of the eastern United States. Golden eagles have caused livestock losses and there have well-been documented cases of golden eagles taking mule deer and pronghorn antelope fawns.
Golden eagle prey consists primarily of small mammals such as prairie dogs, rabbits and ground squirrels, but they will consume birds and reptiles when they can catch them. Large mammals are fair game too. Golden eagles sometimes attack large mammals. Mule deer and pronghorn of all ages have been observed being attacked or killed by eagles. In fact, documented kills of bighorn sheep, coyotes, bobcats, and foxes being killed exist. They also eat carrion.
Many ranchers are not fond of golden eagles. They occasionally kill calves, sheep, pigs and goats. Although attacks on animals that weigh more than about 35 pounds are uncommon, they are not unheard of. The photos below show an adult golden eagle attacking and killing a pronghorn antelope near Rawlins, Wyoming. It was assumed that the pronghorn was not healthy, and the photos are quite gruesome, but they do show the ability of the amazing ability and tenacity of the golden eagle.
As the photos show, golden eagles grab deer and pronghorn anywhere on the head, neck, or body, frequently grasping from the front or side. They kill adult animals with numerous talon stabs into the upper ribs and back. Their feet and talons are well adapted to closing around the backbone, with the talons puncturing large internal arteries, frequently the aorta in front of the kidneys.
The major cause of death is shock produced by massive internal hemorrhage from punctured arteries or collapse of the lungs when the rib cage is punctured. Young animals die from shock and loss of blood as they are eviscerated as golden eagles grab them and simply start eating. Where eagles prey on domestic animals, they usually take the young, but some eagles become persistent predators of livestock as large as 500 pounds. Nature is both cruel and amazing. The golden eagle because of regulations and sound wildlife management practices, but itÂ’s easy to see how this species can have a huge impact livestock producers.
WILDLIFE MANAGEMENT PRO | WILDLIFE AND ENVIRONMENTAL INFO
POSTED UNDER: OUTDOOR NEWS | JANUARY 30
The golden eagle is found across North America and is the largest bird of prey found in the United States. Golden eagles occur in the greatest numbers from Alaska southward throughout the mountain and prairie habitat of the West and into Mexico. They occur in lower numbers to the east across Canada, the Great Lakes states, and the Appalachian Mountains of the eastern United States. Golden eagles have caused livestock losses and there have well-been documented cases of golden eagles taking mule deer and pronghorn antelope fawns.
Golden eagle prey consists primarily of small mammals such as prairie dogs, rabbits and ground squirrels, but they will consume birds and reptiles when they can catch them. Large mammals are fair game too. Golden eagles sometimes attack large mammals. Mule deer and pronghorn of all ages have been observed being attacked or killed by eagles. In fact, documented kills of bighorn sheep, coyotes, bobcats, and foxes being killed exist. They also eat carrion.
Many ranchers are not fond of golden eagles. They occasionally kill calves, sheep, pigs and goats. Although attacks on animals that weigh more than about 35 pounds are uncommon, they are not unheard of. The photos below show an adult golden eagle attacking and killing a pronghorn antelope near Rawlins, Wyoming. It was assumed that the pronghorn was not healthy, and the photos are quite gruesome, but they do show the ability of the amazing ability and tenacity of the golden eagle.
As the photos show, golden eagles grab deer and pronghorn anywhere on the head, neck, or body, frequently grasping from the front or side. They kill adult animals with numerous talon stabs into the upper ribs and back. Their feet and talons are well adapted to closing around the backbone, with the talons puncturing large internal arteries, frequently the aorta in front of the kidneys.
The major cause of death is shock produced by massive internal hemorrhage from punctured arteries or collapse of the lungs when the rib cage is punctured. Young animals die from shock and loss of blood as they are eviscerated as golden eagles grab them and simply start eating. Where eagles prey on domestic animals, they usually take the young, but some eagles become persistent predators of livestock as large as 500 pounds. Nature is both cruel and amazing. The golden eagle because of regulations and sound wildlife management practices, but itÂ’s easy to see how this species can have a huge impact livestock producers.
^ A prime example of the mechanical damage done by the large claws of a bird through stabbing.
What can be made from this?
-Smaller prey specialists like owls and falcons rely on their grip strength for killing rather than their claws (the claws are used to keep the prey from running) while larger prey specialists like eagles and hawks mainly rely on the mechanical damage done by their claws (through stabbing) rather than the pressure exerted from their feet. For smaller prey since Accipitridae lacks the pressure sufficient enough to constrict, they simply use their large claws to hold down smaller prey while they eat it alive.
- Owls and falcons use their beaks to bite down rather than nip on their prey to immobilize the nervous system and prevent escape rather than for killing.
- Due to their foot morphology owls and falcon are incapable of taking large game on the same level as eagle and hawks.
-Due to their foot morphology eagles and hawks are able to tackle much larger prey to compensate for their lacking in the ability of catching smaller fleet footed prey that animals like owls frequently take.
-----------------------------------------------------------------------------------------------------------------------------
SUMMARY
- Accipitridae have longer claws than other birds of prey but in return have weaker grip strengths thus rely more on mechanical damage done by their claws via stabbing (On larger prey) and biting and eating alive (RPR method)
rather than suffocation of prey.
-Smaller prey specialists like owls and falcons rely on their grip strength for killing rather than their claws (the claws are used to keep the prey from running) while larger prey specialists like eagles and hawks mainly rely on the mechanical damage done by their claws (through stabbing) rather than the pressure exerted from their feet. For smaller prey since Accipitridae lacks the pressure sufficient enough to constrict, they simply use their large claws to hold down smaller prey while they eat it alive.
- Owls and falcon have shorter talons but stronger grip strength than eagle and hawks
- Falcon and owl rely more on constriction of prey and nervous system immobilation to kill why acccipitridae rely on mechanical damage via stabbing of large prey or using their large claws for pinning of small prey for live consumption.
Larger prey specialist have larger talons and longer flexor tendon while small prey specialist have it reversed.
- Accipitridae kill larger prey via stabbing of the backbone and head.
- Birds specialized in catching other birds have a medium between grip strength and talon length.
For more info on bird bite force, see here
onlinelibrary.wiley.com/doi/10.1111/j.1365-2435.2008.01494.x/full








