Post by Life on Feb 7, 2021 6:30:20 GMT 5
1. Genus Prognathodon - a case study for the physiology of Mosasaurs
Publication for reference: Soft tissue preservation in a fossil marine lizard with a bilobed tail fin
ABSTRACT
Mosasaurs are secondarily aquatic squamates that became the dominant marine reptiles in the Late Cretaceous about 98–66 million years ago. Although early members of the group possessed body shapes similar to extant monitor lizards, derived forms have traditionally been portrayed as long, sleek animals with broadened, yet ultimately tapering tails. Here we report an extraordinary mosasaur fossil from the Maastrichtian of Harrana in central Jordan, which preserves soft tissues, including high fidelity outlines of a caudal fluke and flippers. This specimen provides the first indisputable evidence that derived mosasaurs were propelled by hypocercal tail fins, a hypothesis that was previously based on comparative skeletal anatomy alone. Ecomorphological comparisons suggest that derived mosasaurs were similar to pelagic sharks in terms of swimming performance, a finding that significantly expands our understanding of the level of aquatic adaptation achieved by these seagoing lizards.
Citation: Lindgren, J., Kaddumi, H. F., & Polcyn, M. J. (2013). Soft tissue preservation in a fossil marine lizard with a bilobed tail fin. Nature Communications, 4(1), 1-8.
Full read: www.nature.com/articles/ncomms3423
---
Figure 3: Prognathodon sp. ERMNH HFV 197-A tail fluke with soft tissues.
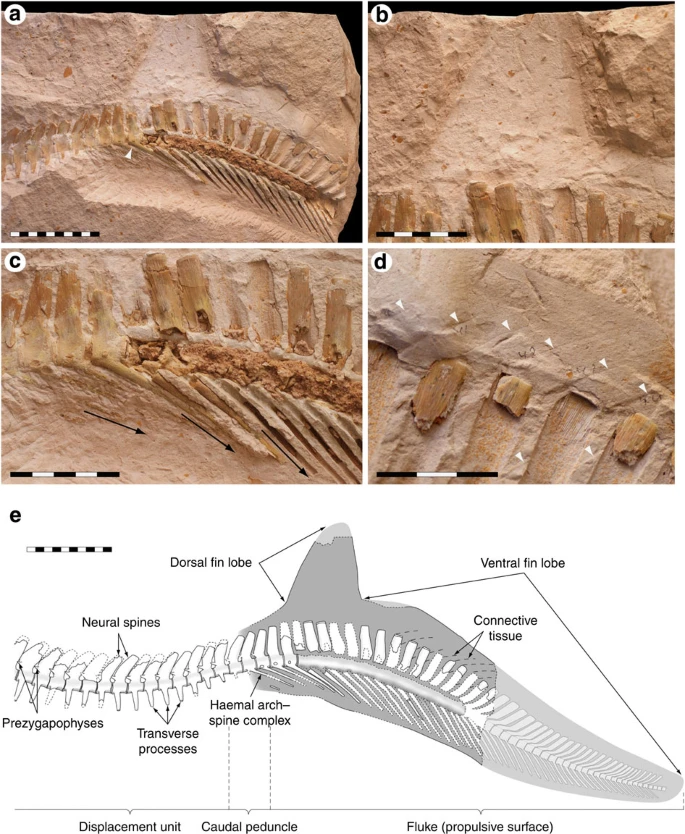
(a) An overview of the caudal region preserving the tail fin. Note that the downturned section of the caudal skeleton lacks transverse processes (the last intermediate caudal is marked with an arrowhead), to suggest that the fluke was laterally flattened. (b) A close-up of the fleshy dorsal fin lobe. Note how the neural spines change in orientation along the depicted vertebral section, forming a vertically dilated, fan-like extremity. (c) Close-up of the vertebrae at the base of the expanded portion of the tail. Arrows indicate the changing inclination of the fused chevrons of the haemal arch–spine complexes at the transition from intermediate to terminal caudal vertebrae, representing their original configuration. (d) Connective tissue (arrowheads) and its relationship to the spinous processes of the vertebrae in the ventral fin lobe. In addition to contributing to a streamlined cross-section, the inclination of the fibrous structures suggests that they served to stiffen the fluke during its sideways excursions. (e) Reconstructed tail fin of Prognathodon sp. Whereas the tip of the dorsal fin lobe was confidently reconstructed from a photograph taken during the excavation of ERMNH HFV 197, the distalmost portion of the tail is based on the caudal skeleton of the closely related mosasaurine Clidastes propython (KUVP 1022; Natural History Museum and Biodiversity Research Center). The number of missing vertebrae was estimated by visual comparison and interpolation of dimensional data. Scale bars, (a, e) 10 cm, (b, c) 5 cm and (d) 3 cm.
---
Figure 6: Reconstructed profile of ERMNH HFV 197 and body shape correlation between taxa.

(a) Hypothetical body outline of Prognathodon based on ERMNH HFV 197 with added data/measurements from P. overtoni and P. saturator. Note that the artificial dorsal tail curvature produced by the pygal and anterior intermediate caudal vertebrae in ERMNH HFV 197 has been eliminated (based on a comparison with the virtually intact and articulated tail of LACM 128319 (refs 22, 52)), resulting in a deeper caudal fin with a higher aspect ratio. (b) Body and tail form distribution in modern scyliorhinid, carcharhinid and lamnid sharks, two Mesozoic ichthyosaurs (Chensaurus and Stenopterygius) and Prognathodon. The diagram in is redrawn from ref. 55 with added data from ERMNH HFV 197.
-----
2. Evolution
Publication for reference: Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas
Abstract
Observations of temporal overlap of niche occupation among Late Cretaceous marine amniotes suggest that the rise and diversification of mosasauroid squamates might have been influenced by competition with or disappearance of some plesiosaur taxa. We discuss that hypothesis through comparisons of the rates of morphological evolution of mosasauroids throughout their evolutionary history with those inferred for contemporary plesiosaur clades. We used expanded versions of two species-level phylogenetic datasets of both these groups, updated them with stratigraphic information, and analyzed using the Bayesian inference to estimate the rates of divergence for each clade. The oscillations in evolutionary rates of the mosasauroid and plesiosaur lineages that overlapped in time and space were then used as a baseline for discussion and comparisons of traits that can affect the shape of the niche structures of aquatic amniotes, such as tooth morphologies, body size, swimming abilities, metabolism, and reproduction. Only two groups of plesiosaurs are considered to be possible niche competitors of mosasauroids: the brachauchenine pliosaurids and the polycotylid leptocleidians. However, direct evidence for interactions between mosasauroids and plesiosaurs is scarce and limited only to large mosasauroids as the predators/scavengers and polycotylids as their prey. The first mosasauroids differed from contemporary plesiosaurs in certain aspects of all discussed traits and no evidence suggests that early representatives of Mosasauroidea diversified after competitions with plesiosaurs. Nevertheless, some mosasauroids, such as tylosaurines, might have seized the opportunity and occupied the niche previously inhabited by brachauchenines, around or immediately after they became extinct, and by polycotylids that decreased their phylogenetic diversity and disparity around the time the large-sized tylosaurines started to flourish.
Citation: Madzia, D., & Cau, A. (2020). Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas. PeerJ, 8, e8941.
Full read: www.ncbi.nlm.nih.gov/pmc/articles/PMC7164395/
---
Figure 1: Maximum Clade Credibility Tree (MCCT) of Mosasauroidea.

MCCT of Mosasauroidea, rooted on Adriosaurus. Branches colored according to median rate of divergence. Time scale in My. Values above branches (bold) indicate the posterior probability of the clade; values at nodes indicate median age of divergence of node (Ma). Circles on nodes indicate areas of origin reconstructed for major mosasauroid clades.
---
Findings:
-----
3. Pathologies - Evidence of trophic interactions and susceptibility to bone disease
Publication for reference: Co-Ossification of Vertebrae in Mosasaurs (Squamata, Mosasauridae); Evidence of Habitat Interactions and Susceptibility to Bone Disease
Abstract
Pathologies involving the fusion of mosasaur vertebrae have been documented in the literature since the late 1870s. Although some instances can clearly be demonstrated to be the result of an attack by another predator, usually a shark, the source of the pathology is more difficult to discern in other specimens. Here we discuss the evidence for pathologies as the result of interactions with the mosasaur's habitat and the apparent susceptibility of certain kinds of disease affecting the skeleton as preserved in the specimens.
Citation: Rothschild, B., & Everhart, M. J. (2015). Co-ossification of vertebrae in mosasaurs (Squamata, Mosasauridae); Evidence of habitat interactions and susceptibility to bone disease. Transactions of the Kansas Academy of Science, 118(3-4), 265-275.
LINK: bioone.org/journals/transactions-of-the-kansas-academy-of-science/volume-118/issue-3-4/062.118.0309/Co-Ossification-of-Vertebrae-in-Mosasaurs-Squamata-Mosasauridae-Evidence-of/10.1660/062.118.0309.short
---

Figure 1. A. Adult Platecarpus tympaniticus (FHSM VP-322) caudal vertebrae with two widely separated vertebral fusions (arrows). Scale bar = 10 cm. B. Fused caudal vertebrae at the end of the tail of a “club tail” Tylosaurus (FHSM VP-13750) in left lateral view (upper) and right lateral view (lower). Anterior is to the left. The embedded tooth of a shark, the apparent cause of the infection, is located within the white oval. After healing, the fused mass and 28 associated caudal vertebrae of this specimen were subsequently consumed and partially digested by another predator. Scale bar = 2 cm. C. the fusion of three caudal vertebrae near the end of the tail of another Tylosaurus specimen (FHSM VP-17984) with a “club tail.” Scale bar = 5 cm.
---
Findings:

-----
4. Calcium isotopic analysis - a case study for the extinction of Mosasaurs
Publication for reference: Calcium Isotopic Evidence for Vulnerable Marine Ecosystem Structure Prior to the K/Pg Extinction
SUMMARY
The collapse of marine ecosystems during the end-Cretaceous mass extinction involved the base of the food chain [1] up to ubiquitous vertebrate apex predators [2, 3, 4, 5]. Large marine reptiles became suddenly extinct at the Cretaceous-Paleogene (K/Pg) boundary, whereas other contemporaneous groups such as bothremydid turtles or dyrosaurid crocodylomorphs, although affected at the familial, genus, or species level, survived into post-crisis environments of the Paleocene [5, 6, 7, 8, 9] and could have found refuge in freshwater habitats [10, 11, 12]. A recent hypothesis proposes that the extinction of plesiosaurians and mosasaurids could have been caused by an important drop in sea level [13]. Mosasaurids are unusually diverse and locally abundant in the Maastrichtian phosphatic deposits of Morocco, and with large sharks and one species of elasmosaurid plesiosaurian recognized so far, contribute to an overabundance of apex predators [3, 7, 14, 15]. For this reason, high local diversity of marine reptiles exhibiting different body masses and a wealth of tooth morphologies hints at complex trophic interactions within this latest Cretaceous marine ecosystem. Using calcium isotopes, we investigated the trophic structure of this extinct assemblage. Our results are consistent with a calcium isotope pattern observed in modern marine ecosystems and show that plesiosaurians and mosasaurids indiscriminately fall in the tertiary piscivore group. This suggests that marine reptile apex predators relied onto a single dietary calcium source, compatible with the vulnerable wasp-waist food webs of the modern world [16]. This inferred peculiar ecosystem structure may help explain plesiosaurian and mosasaurid extinction following the end-Cretaceous biological crisis.
Citation: Martin, J. E., Vincent, P., Tacail, T., Khaldoune, F., Jourani, E., Bardet, N., & Balter, V. (2017). Calcium isotopic evidence for vulnerable marine ecosystem structure prior to the K/Pg extinction. Current Biology, 27(11), 1641-1644.
Full read: www.sciencedirect.com/science/article/pii/S0960982217304876#bib16
-----
Findings:

Figure 1. δ44/42Ca Variability in Marine Ecosystems of the Modern World and the Maastrichtian
δ44/42Ca data are given in ‰ relative to ICP Ca Lyon standard. For the modern dataset (A), data compilation is from the literature (see [19] for details), with the exception of modern marine turtles (this study). The fossil Maastrichtian dataset (B) is entirely new to this study. 1, modern marine turtles; 2, zooplanktivores; 3, primary piscivores; 4, secondary piscivores; 5, tertiary piscivores; 7, Maastrichtian marine turtles; 8, Maastrichtian primary piscivores; 9, Maastrichtian tertiary piscivores; 10, mosasaurs; 11, plesiosaurs. Triangles represent invertebrates; circles represent marine reptiles; squares represent teleost fishes; diamonds represent elasmobranchs. Error bars represent 2SD. See also Table S1.
-----
Extinction hypothesis:
-----
Publication for reference: Soft tissue preservation in a fossil marine lizard with a bilobed tail fin
ABSTRACT
Mosasaurs are secondarily aquatic squamates that became the dominant marine reptiles in the Late Cretaceous about 98–66 million years ago. Although early members of the group possessed body shapes similar to extant monitor lizards, derived forms have traditionally been portrayed as long, sleek animals with broadened, yet ultimately tapering tails. Here we report an extraordinary mosasaur fossil from the Maastrichtian of Harrana in central Jordan, which preserves soft tissues, including high fidelity outlines of a caudal fluke and flippers. This specimen provides the first indisputable evidence that derived mosasaurs were propelled by hypocercal tail fins, a hypothesis that was previously based on comparative skeletal anatomy alone. Ecomorphological comparisons suggest that derived mosasaurs were similar to pelagic sharks in terms of swimming performance, a finding that significantly expands our understanding of the level of aquatic adaptation achieved by these seagoing lizards.
Citation: Lindgren, J., Kaddumi, H. F., & Polcyn, M. J. (2013). Soft tissue preservation in a fossil marine lizard with a bilobed tail fin. Nature Communications, 4(1), 1-8.
Full read: www.nature.com/articles/ncomms3423
---
Figure 3: Prognathodon sp. ERMNH HFV 197-A tail fluke with soft tissues.
(a) An overview of the caudal region preserving the tail fin. Note that the downturned section of the caudal skeleton lacks transverse processes (the last intermediate caudal is marked with an arrowhead), to suggest that the fluke was laterally flattened. (b) A close-up of the fleshy dorsal fin lobe. Note how the neural spines change in orientation along the depicted vertebral section, forming a vertically dilated, fan-like extremity. (c) Close-up of the vertebrae at the base of the expanded portion of the tail. Arrows indicate the changing inclination of the fused chevrons of the haemal arch–spine complexes at the transition from intermediate to terminal caudal vertebrae, representing their original configuration. (d) Connective tissue (arrowheads) and its relationship to the spinous processes of the vertebrae in the ventral fin lobe. In addition to contributing to a streamlined cross-section, the inclination of the fibrous structures suggests that they served to stiffen the fluke during its sideways excursions. (e) Reconstructed tail fin of Prognathodon sp. Whereas the tip of the dorsal fin lobe was confidently reconstructed from a photograph taken during the excavation of ERMNH HFV 197, the distalmost portion of the tail is based on the caudal skeleton of the closely related mosasaurine Clidastes propython (KUVP 1022; Natural History Museum and Biodiversity Research Center). The number of missing vertebrae was estimated by visual comparison and interpolation of dimensional data. Scale bars, (a, e) 10 cm, (b, c) 5 cm and (d) 3 cm.
---
Figure 6: Reconstructed profile of ERMNH HFV 197 and body shape correlation between taxa.
(a) Hypothetical body outline of Prognathodon based on ERMNH HFV 197 with added data/measurements from P. overtoni and P. saturator. Note that the artificial dorsal tail curvature produced by the pygal and anterior intermediate caudal vertebrae in ERMNH HFV 197 has been eliminated (based on a comparison with the virtually intact and articulated tail of LACM 128319 (refs 22, 52)), resulting in a deeper caudal fin with a higher aspect ratio. (b) Body and tail form distribution in modern scyliorhinid, carcharhinid and lamnid sharks, two Mesozoic ichthyosaurs (Chensaurus and Stenopterygius) and Prognathodon. The diagram in is redrawn from ref. 55 with added data from ERMNH HFV 197.
-----
2. Evolution
Publication for reference: Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas
Abstract
Observations of temporal overlap of niche occupation among Late Cretaceous marine amniotes suggest that the rise and diversification of mosasauroid squamates might have been influenced by competition with or disappearance of some plesiosaur taxa. We discuss that hypothesis through comparisons of the rates of morphological evolution of mosasauroids throughout their evolutionary history with those inferred for contemporary plesiosaur clades. We used expanded versions of two species-level phylogenetic datasets of both these groups, updated them with stratigraphic information, and analyzed using the Bayesian inference to estimate the rates of divergence for each clade. The oscillations in evolutionary rates of the mosasauroid and plesiosaur lineages that overlapped in time and space were then used as a baseline for discussion and comparisons of traits that can affect the shape of the niche structures of aquatic amniotes, such as tooth morphologies, body size, swimming abilities, metabolism, and reproduction. Only two groups of plesiosaurs are considered to be possible niche competitors of mosasauroids: the brachauchenine pliosaurids and the polycotylid leptocleidians. However, direct evidence for interactions between mosasauroids and plesiosaurs is scarce and limited only to large mosasauroids as the predators/scavengers and polycotylids as their prey. The first mosasauroids differed from contemporary plesiosaurs in certain aspects of all discussed traits and no evidence suggests that early representatives of Mosasauroidea diversified after competitions with plesiosaurs. Nevertheless, some mosasauroids, such as tylosaurines, might have seized the opportunity and occupied the niche previously inhabited by brachauchenines, around or immediately after they became extinct, and by polycotylids that decreased their phylogenetic diversity and disparity around the time the large-sized tylosaurines started to flourish.
Citation: Madzia, D., & Cau, A. (2020). Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas. PeerJ, 8, e8941.
Full read: www.ncbi.nlm.nih.gov/pmc/articles/PMC7164395/
---
Figure 1: Maximum Clade Credibility Tree (MCCT) of Mosasauroidea.
MCCT of Mosasauroidea, rooted on Adriosaurus. Branches colored according to median rate of divergence. Time scale in My. Values above branches (bold) indicate the posterior probability of the clade; values at nodes indicate median age of divergence of node (Ma). Circles on nodes indicate areas of origin reconstructed for major mosasauroid clades.
---
Findings:
Body size evolution
The earliest mosasauroids, such as Aigialosaurus, Carsosaurus, Haasiasaurus, Komensaurus, and Opetiosaurus, had slim and elongated bodies, with a total length of about 1 to 2 m, which strongly contrasts with the bulky, multitone contemporary plesiosaurs. However, larger-sized mosasauroids appeared relative early in the evolutionary history of the clade, as is documented, for example, by the specimen TMM 43345-1, which represents a large tylosaurine (Bell, Barnes & Polcyn, 2013). The specimen originates from the upper middle Turonian of the Ernst Member (Boquillas Formation) of Texas; thus, tylosaurines evolved larger-sized forms around the time or shortly after brachauchenines became extinct. Nevertheless, members of other mosasauroid clades (plioplatecarpines and mosasaurines) evolved large sizes (>5 m in length) relatively early as well (see, e.g., Polcyn et al., 2014: Appendix A. Supplementary data).
Evolution of swimming abilities
Swimming modes and abilities of Mesozoic aquatic vertebrates are tightly connected with their physiology, behavior, and other aspects of their biology and, thus, constitute an important research area (e.g., Massare, 1988; Massare, 1994; Motani, 2002). A number of studies have thoroughly assessed the swimming abilities in plesiosaurs and discussed the differences in the two main plesiosaur ‘body plans’ –the ‘long-’ and ‘short-necked’ forms (e.g., Frey & Riess, 1982; Tarsitano & Riess, 1982; Godfrey, 1984; Halstead, 1989; Nicholls & Russell, 1991; Lingham-Soliar, 2000; O’Keefe, 2001; Carpenter et al., 2010; Liu et al., 2015; Muscutt et al., 2017; Noè, Taylor & Gómez-Pérez, 2017; Troelsen et al., 2019). The same applies to mosasauroids whose swimming abilities and especially their origin have been assessed through detailed studies of various aspects of their anatomy (see, e.g., Lindgren, Jagt & Caldwell, 2007; Lindgren et al., 2010; Lindgren, Polcyn & Young, 2011; Konishi et al., 2012; LeBlanc, Caldwell & Lindgren, 2013; Lindgren, Kaddumi & Polcyn, 2013; Houssaye & Bardet, 2013; Cuthbertson et al., 2015; D’Emic, Smith & Ansley, 2015). The mode of swimming in the two clades is known to have differed greatly. Plesiosaurs have exhibited a limb-based propulsion while mosasauroids employed lateral undulatory locomotion. These differences also apparently reflect the modes of predation in these groups. Owing to their anatomical similarities—large heads, relatively short necks, bulky bodies, and rather short tails—brachauchenines and polycotylids were specialized for maneuverability and pursuit (e.g., O’Keefe, 2001). Mosasauroids, in turn, had long been characterized as being slower-swimming predators adapted for brief ambush pursuits (e.g., Massare, 1988; Massare, 1994; Motani, 2002). Over the last few years, however, the knowledge of the mosasauroid body plan evolution has improved considerably (see, e.g., discussions in Lindgren, Polcyn & Young (2011) and Lindgren, Kaddumi & Polcyn (2013), suggesting that the swimming performance of derived mosasauroids was similar to that of pelagic sharks.
In general, mosasauroids comprised a wide array of taxa; from semi-aquatic forms (e.g., Bell & Polcyn, 2005; Polcyn & Bell, 2005; Dutchak & Caldwell, 2006; Dutchak & Caldwell, 2009; Caldwell & Palci, 2007; Mekarski et al., 2019) to fully aquatic swimmers (see, e.g., Lindgren, Jagt & Caldwell, 2007). Nevertheless, the course and timing of their transition from semi- to fully aquatic morphologies (that is, from ‘plesiopedal-plesiopelvic’ to ‘hydropedal-hydropelvic’ conditions; sensu Bell & Polcyn (2005) and Caldwell & Palci (2007), a key aspect when considering potential competitive interactions between mosasauroids and plesiosaurs, is somewhat hindered by conflicting hypotheses of the early evolution of the group. Current phylogenetic assessments of the mosasauroid basal branching are highly dependable on the tree-search strategies used (Simões et al., 2017; Madzia & Cau, 2017). For instance, out of several phylogenetic methods applied, only the parsimony analysis with implied weighting performed by Simões et al. (2017), with the default setting of the concavity parameter (K = 3), inferred a single origin of the fully aquatic lifestyle in mosasauroids (with a reversal to the ‘plesiopelvic’ condition in tethysaurines). Madzia & Cau (2017) questioned these findings on the ground of the ongoing debate regarding the meaning of the K parameter (O’Reilly et al., 2016; Congreve & Lamsdell, 2016; Goloboff, Torres & Arias, 2018) and also the lack of multiple approaches to the phylogenetic assessment using the implied weighting function (see Goloboff, 1993; Goloboff, 1995; Goloboff et al., 2008; Goloboff, Torres & Arias, 2018). It is also essential to note that the use of the K-value that is set as default in TNT (that is, K = 3), appears to be too strong and leading to unnatural grouping of OTUs, especially for larger datasets (Goloboff, Torres & Arias, 2018). Thus, higher (though not too high) values should be preferred (see also discussion in Herne et al., 2019: Supplemental text S1: 9–12]).
Considering the results stemming from the most recent parsimony and Bayesian analyses (Simões et al., 2017; Madzia & Cau, 2017; this study), mosasauroids might have evolved the fully aquatic lifestyle more than once. Still, the course of the transition remains a subject for detailed multidisciplinary assessments. For example, despite that the study of Houssaye et al. (2013) was focused on mosasaurine mosasauroids, the authors analyzed the limb-bone osteohistology in a wide variety of taxa, including the basal mosasaurine Dallasaurus, a taxon with a ‘plesiopedal’/‘hydropelvic’ morphology that is ‘transitional’ between semi- and fully-aquatic forms, as well as specimens assigned to the fully aquatic (‘hydropedal’/‘hydropelvic’) taxa Clidastes, Globidens, Mosasaurus, Plotosaurus, and ‘Prognathodon’. The results, when further compared with previous osteohistological studies (e.g., Houssaye & Bardet, 2013), revealed that ‘transitional’ mosasauroids, or at least those representing forms intermediate between basal semi-aquatic mosasauroids and advanced fully-aquatic mosasaurines, exhibited a peculiar inner bone organization characterized by the combination of terrestrial-like and aquatic features that suggested a more gradual adaptation to open marine environments than previously thought. Interestingly, the acquisition of the ‘hydropedal’ and ‘hydropelvic’ conditions, as inferred through our Bayesian analysis, occurred approximately at the time and within the lineages with the highest rates of evolution, resulting in the appearance of good swimmers around the time the brachauchenines experienced low rates of morphological evolution and died out.
Thermoregulation and metabolic rates
Oxygen isotope compositions (δ18O) data obtained from the tooth phosphate of plesiosaurs suggest that they were able to regulate their body temperature independently of the surrounding waters and had high metabolic rates that are required for fast swimming over large distances and predation (Bernard et al., 2010). Plesiosaur metabolic rates have been later independently assessed through the study of their osteohistology (Fleischle, Wintrich & Sander, 2018), which supported the inference of high rates in the clade. In mosasauroids, the available evidence offers slightly ambiguous results. Bernard et al. (2010) proposed that the body temperature of mosasauroids could have been at least partly influenced by ambient conditions; still, they found support for high metabolic rates in mosasauroids. While reassessing the results of Bernard et al. (2010) and Motani (2010) noted that the temperatures provided in Bernard et al. (2010) might be artifacts arising from time-dependent depletion of δ18O (see also Veizer, Godderis & François, 2000), and argued that mosasauroids might have had a tendency to overheat, proposing that they may have been gigantothermic. Nevertheless, such conclusions appear to be in disagreement with a further stable oxygen isotope study (Harrell, Pérez-Huerta & Suarez, 2016) that characterized mosasauroids as being endotherms rather than gigantotherms. With respect to the mosasauroid metabolic rates, Houssaye et al.’s (2013) osteohistological study showed that their basal metabolic rates were intermediate between those of the extant leatherback turtles (that are homeothermic but not endothermic; e.g., Motani (2010) and Houssaye (2013) and those inferred for plesiosaurs (that are endothermic e.g., Bernard et al., 2010; Motani, 2010; Houssaye, 2013; Fleischle, Wintrich & Sander, 2018; Fleischle et al., 2019).
Reproduction and early life history
Available evidence related to the reproductive strategies and early life history in mosasauroids and plesiosaurs is currently limited to a few studies (e.g., Caldwell & Lee, 2001; Kear, 2007; O’Keefe & Chiappe, 2011; Houssaye & Tafforeau, 2012; Houssaye & Bardet, 2013; Field et al., 2015) and reports that have not been published beyond conference abstracts (e.g., Bell et al., 1996; Everhart, 2002; Bell & Sheldon, 2004). A study describing the first gravid plesiosaur, a polycotylid specimen referred to Polycotylus latipinnis (O’Keefe & Chiappe, 2011), has initiated comparisons between reproductive strategies of plesiosaurs and other marine amniotes, including mosasauroids. Both clades, mosasauroids and plesiosaurs, have been viviparous though their reproductive strategies differed. In the early (semi-aquatic) mosasauroid Carsosaurus, females have been apparently giving birth to at least four progenies (Caldwell & Lee, 2001). Published record does not provide definitive answer regarding the number of embryos in more advanced (larger and fully aquatic) members of Mosasauroidea though preliminary reports suggested that plioplatecarpines were giving birth to multiple progenies as well (Bell & Sheldon, 2004). The gravid plesiosaur specimen, in turn, shows only a single fetus (O’Keefe & Chiappe, 2011). Following comparisons of the traits observed in that specimen to those in the closest extant ecological analogs (odontocete cetaceans) and taxa with some plesiosaur-like reproductive traits (Egernia spp.), O’Keefe & Chiappe (2011) suggested that plesiosaurs were K-selected and hypothesized that they were social and may have been engaged in parental care. It could be speculated that multiple progenies in mosasauroids, if also present in large-sized forms, might have given these squamates some advantages over plesiosaurs, especially if they were born in open pelagic setting and immediately occupied it (e.g., Houssaye & Tafforeau, 2012; Field et al., 2015). When such things are considered, it is worth noting, however, that the theory of r/K-selection of MacArthur & Wilson (1967), a paradigm popular as a predictive model for life-history evolution in the late 1960s and 1970s (see also Pianka, 1970), has long been challenged (see, e.g., discussion in Reznick, Bryant & Bashey, 2002). Even if mosasauroids and plesiosaurs differed in both, their reproductive strategies and early life history, the evolutionary meaning of these differences and their impact on the life history of mosasauroids and plesiosaurs, when assessed from the perspective of their niche occupation, remains unknown.
The earliest mosasauroids, such as Aigialosaurus, Carsosaurus, Haasiasaurus, Komensaurus, and Opetiosaurus, had slim and elongated bodies, with a total length of about 1 to 2 m, which strongly contrasts with the bulky, multitone contemporary plesiosaurs. However, larger-sized mosasauroids appeared relative early in the evolutionary history of the clade, as is documented, for example, by the specimen TMM 43345-1, which represents a large tylosaurine (Bell, Barnes & Polcyn, 2013). The specimen originates from the upper middle Turonian of the Ernst Member (Boquillas Formation) of Texas; thus, tylosaurines evolved larger-sized forms around the time or shortly after brachauchenines became extinct. Nevertheless, members of other mosasauroid clades (plioplatecarpines and mosasaurines) evolved large sizes (>5 m in length) relatively early as well (see, e.g., Polcyn et al., 2014: Appendix A. Supplementary data).
Evolution of swimming abilities
Swimming modes and abilities of Mesozoic aquatic vertebrates are tightly connected with their physiology, behavior, and other aspects of their biology and, thus, constitute an important research area (e.g., Massare, 1988; Massare, 1994; Motani, 2002). A number of studies have thoroughly assessed the swimming abilities in plesiosaurs and discussed the differences in the two main plesiosaur ‘body plans’ –the ‘long-’ and ‘short-necked’ forms (e.g., Frey & Riess, 1982; Tarsitano & Riess, 1982; Godfrey, 1984; Halstead, 1989; Nicholls & Russell, 1991; Lingham-Soliar, 2000; O’Keefe, 2001; Carpenter et al., 2010; Liu et al., 2015; Muscutt et al., 2017; Noè, Taylor & Gómez-Pérez, 2017; Troelsen et al., 2019). The same applies to mosasauroids whose swimming abilities and especially their origin have been assessed through detailed studies of various aspects of their anatomy (see, e.g., Lindgren, Jagt & Caldwell, 2007; Lindgren et al., 2010; Lindgren, Polcyn & Young, 2011; Konishi et al., 2012; LeBlanc, Caldwell & Lindgren, 2013; Lindgren, Kaddumi & Polcyn, 2013; Houssaye & Bardet, 2013; Cuthbertson et al., 2015; D’Emic, Smith & Ansley, 2015). The mode of swimming in the two clades is known to have differed greatly. Plesiosaurs have exhibited a limb-based propulsion while mosasauroids employed lateral undulatory locomotion. These differences also apparently reflect the modes of predation in these groups. Owing to their anatomical similarities—large heads, relatively short necks, bulky bodies, and rather short tails—brachauchenines and polycotylids were specialized for maneuverability and pursuit (e.g., O’Keefe, 2001). Mosasauroids, in turn, had long been characterized as being slower-swimming predators adapted for brief ambush pursuits (e.g., Massare, 1988; Massare, 1994; Motani, 2002). Over the last few years, however, the knowledge of the mosasauroid body plan evolution has improved considerably (see, e.g., discussions in Lindgren, Polcyn & Young (2011) and Lindgren, Kaddumi & Polcyn (2013), suggesting that the swimming performance of derived mosasauroids was similar to that of pelagic sharks.
In general, mosasauroids comprised a wide array of taxa; from semi-aquatic forms (e.g., Bell & Polcyn, 2005; Polcyn & Bell, 2005; Dutchak & Caldwell, 2006; Dutchak & Caldwell, 2009; Caldwell & Palci, 2007; Mekarski et al., 2019) to fully aquatic swimmers (see, e.g., Lindgren, Jagt & Caldwell, 2007). Nevertheless, the course and timing of their transition from semi- to fully aquatic morphologies (that is, from ‘plesiopedal-plesiopelvic’ to ‘hydropedal-hydropelvic’ conditions; sensu Bell & Polcyn (2005) and Caldwell & Palci (2007), a key aspect when considering potential competitive interactions between mosasauroids and plesiosaurs, is somewhat hindered by conflicting hypotheses of the early evolution of the group. Current phylogenetic assessments of the mosasauroid basal branching are highly dependable on the tree-search strategies used (Simões et al., 2017; Madzia & Cau, 2017). For instance, out of several phylogenetic methods applied, only the parsimony analysis with implied weighting performed by Simões et al. (2017), with the default setting of the concavity parameter (K = 3), inferred a single origin of the fully aquatic lifestyle in mosasauroids (with a reversal to the ‘plesiopelvic’ condition in tethysaurines). Madzia & Cau (2017) questioned these findings on the ground of the ongoing debate regarding the meaning of the K parameter (O’Reilly et al., 2016; Congreve & Lamsdell, 2016; Goloboff, Torres & Arias, 2018) and also the lack of multiple approaches to the phylogenetic assessment using the implied weighting function (see Goloboff, 1993; Goloboff, 1995; Goloboff et al., 2008; Goloboff, Torres & Arias, 2018). It is also essential to note that the use of the K-value that is set as default in TNT (that is, K = 3), appears to be too strong and leading to unnatural grouping of OTUs, especially for larger datasets (Goloboff, Torres & Arias, 2018). Thus, higher (though not too high) values should be preferred (see also discussion in Herne et al., 2019: Supplemental text S1: 9–12]).
Considering the results stemming from the most recent parsimony and Bayesian analyses (Simões et al., 2017; Madzia & Cau, 2017; this study), mosasauroids might have evolved the fully aquatic lifestyle more than once. Still, the course of the transition remains a subject for detailed multidisciplinary assessments. For example, despite that the study of Houssaye et al. (2013) was focused on mosasaurine mosasauroids, the authors analyzed the limb-bone osteohistology in a wide variety of taxa, including the basal mosasaurine Dallasaurus, a taxon with a ‘plesiopedal’/‘hydropelvic’ morphology that is ‘transitional’ between semi- and fully-aquatic forms, as well as specimens assigned to the fully aquatic (‘hydropedal’/‘hydropelvic’) taxa Clidastes, Globidens, Mosasaurus, Plotosaurus, and ‘Prognathodon’. The results, when further compared with previous osteohistological studies (e.g., Houssaye & Bardet, 2013), revealed that ‘transitional’ mosasauroids, or at least those representing forms intermediate between basal semi-aquatic mosasauroids and advanced fully-aquatic mosasaurines, exhibited a peculiar inner bone organization characterized by the combination of terrestrial-like and aquatic features that suggested a more gradual adaptation to open marine environments than previously thought. Interestingly, the acquisition of the ‘hydropedal’ and ‘hydropelvic’ conditions, as inferred through our Bayesian analysis, occurred approximately at the time and within the lineages with the highest rates of evolution, resulting in the appearance of good swimmers around the time the brachauchenines experienced low rates of morphological evolution and died out.
Thermoregulation and metabolic rates
Oxygen isotope compositions (δ18O) data obtained from the tooth phosphate of plesiosaurs suggest that they were able to regulate their body temperature independently of the surrounding waters and had high metabolic rates that are required for fast swimming over large distances and predation (Bernard et al., 2010). Plesiosaur metabolic rates have been later independently assessed through the study of their osteohistology (Fleischle, Wintrich & Sander, 2018), which supported the inference of high rates in the clade. In mosasauroids, the available evidence offers slightly ambiguous results. Bernard et al. (2010) proposed that the body temperature of mosasauroids could have been at least partly influenced by ambient conditions; still, they found support for high metabolic rates in mosasauroids. While reassessing the results of Bernard et al. (2010) and Motani (2010) noted that the temperatures provided in Bernard et al. (2010) might be artifacts arising from time-dependent depletion of δ18O (see also Veizer, Godderis & François, 2000), and argued that mosasauroids might have had a tendency to overheat, proposing that they may have been gigantothermic. Nevertheless, such conclusions appear to be in disagreement with a further stable oxygen isotope study (Harrell, Pérez-Huerta & Suarez, 2016) that characterized mosasauroids as being endotherms rather than gigantotherms. With respect to the mosasauroid metabolic rates, Houssaye et al.’s (2013) osteohistological study showed that their basal metabolic rates were intermediate between those of the extant leatherback turtles (that are homeothermic but not endothermic; e.g., Motani (2010) and Houssaye (2013) and those inferred for plesiosaurs (that are endothermic e.g., Bernard et al., 2010; Motani, 2010; Houssaye, 2013; Fleischle, Wintrich & Sander, 2018; Fleischle et al., 2019).
Reproduction and early life history
Available evidence related to the reproductive strategies and early life history in mosasauroids and plesiosaurs is currently limited to a few studies (e.g., Caldwell & Lee, 2001; Kear, 2007; O’Keefe & Chiappe, 2011; Houssaye & Tafforeau, 2012; Houssaye & Bardet, 2013; Field et al., 2015) and reports that have not been published beyond conference abstracts (e.g., Bell et al., 1996; Everhart, 2002; Bell & Sheldon, 2004). A study describing the first gravid plesiosaur, a polycotylid specimen referred to Polycotylus latipinnis (O’Keefe & Chiappe, 2011), has initiated comparisons between reproductive strategies of plesiosaurs and other marine amniotes, including mosasauroids. Both clades, mosasauroids and plesiosaurs, have been viviparous though their reproductive strategies differed. In the early (semi-aquatic) mosasauroid Carsosaurus, females have been apparently giving birth to at least four progenies (Caldwell & Lee, 2001). Published record does not provide definitive answer regarding the number of embryos in more advanced (larger and fully aquatic) members of Mosasauroidea though preliminary reports suggested that plioplatecarpines were giving birth to multiple progenies as well (Bell & Sheldon, 2004). The gravid plesiosaur specimen, in turn, shows only a single fetus (O’Keefe & Chiappe, 2011). Following comparisons of the traits observed in that specimen to those in the closest extant ecological analogs (odontocete cetaceans) and taxa with some plesiosaur-like reproductive traits (Egernia spp.), O’Keefe & Chiappe (2011) suggested that plesiosaurs were K-selected and hypothesized that they were social and may have been engaged in parental care. It could be speculated that multiple progenies in mosasauroids, if also present in large-sized forms, might have given these squamates some advantages over plesiosaurs, especially if they were born in open pelagic setting and immediately occupied it (e.g., Houssaye & Tafforeau, 2012; Field et al., 2015). When such things are considered, it is worth noting, however, that the theory of r/K-selection of MacArthur & Wilson (1967), a paradigm popular as a predictive model for life-history evolution in the late 1960s and 1970s (see also Pianka, 1970), has long been challenged (see, e.g., discussion in Reznick, Bryant & Bashey, 2002). Even if mosasauroids and plesiosaurs differed in both, their reproductive strategies and early life history, the evolutionary meaning of these differences and their impact on the life history of mosasauroids and plesiosaurs, when assessed from the perspective of their niche occupation, remains unknown.
-----
3. Pathologies - Evidence of trophic interactions and susceptibility to bone disease
Publication for reference: Co-Ossification of Vertebrae in Mosasaurs (Squamata, Mosasauridae); Evidence of Habitat Interactions and Susceptibility to Bone Disease
Abstract
Pathologies involving the fusion of mosasaur vertebrae have been documented in the literature since the late 1870s. Although some instances can clearly be demonstrated to be the result of an attack by another predator, usually a shark, the source of the pathology is more difficult to discern in other specimens. Here we discuss the evidence for pathologies as the result of interactions with the mosasaur's habitat and the apparent susceptibility of certain kinds of disease affecting the skeleton as preserved in the specimens.
Citation: Rothschild, B., & Everhart, M. J. (2015). Co-ossification of vertebrae in mosasaurs (Squamata, Mosasauridae); Evidence of habitat interactions and susceptibility to bone disease. Transactions of the Kansas Academy of Science, 118(3-4), 265-275.
LINK: bioone.org/journals/transactions-of-the-kansas-academy-of-science/volume-118/issue-3-4/062.118.0309/Co-Ossification-of-Vertebrae-in-Mosasaurs-Squamata-Mosasauridae-Evidence-of/10.1660/062.118.0309.short
---
Figure 1. A. Adult Platecarpus tympaniticus (FHSM VP-322) caudal vertebrae with two widely separated vertebral fusions (arrows). Scale bar = 10 cm. B. Fused caudal vertebrae at the end of the tail of a “club tail” Tylosaurus (FHSM VP-13750) in left lateral view (upper) and right lateral view (lower). Anterior is to the left. The embedded tooth of a shark, the apparent cause of the infection, is located within the white oval. After healing, the fused mass and 28 associated caudal vertebrae of this specimen were subsequently consumed and partially digested by another predator. Scale bar = 2 cm. C. the fusion of three caudal vertebrae near the end of the tail of another Tylosaurus specimen (FHSM VP-17984) with a “club tail.” Scale bar = 5 cm.
---
Findings:
A residual question is why spondyloarthropathy was more common in Plioplatecarpus and Clidastes than in Platecarpus, Tylosaurus and Mosasaurus? Spondyloarthropathy is a category of arthritis that consists of 5 varieties: The skin disease psoriasis, inflammatory bowel disease and ankylosing spondylitis, reactive arthritis, and an undifferentiated arthritis (Resnick 2002; Rothschild and Martin 2006). Ankylosing spondylitis starts at the pelvis (lumbar/caudal junction) and progresses caudally, a different pattern than that noted in mosasaurs. Skin pathology has yet to be reported, but resolving this possibility would require detailed evaluation of skin impressions by a dermatologist. Inflammatory bowel disease could be considered but a reactive or undifferentiated form of arthritis appears more likely, especially in view of possible environmental implications. Gastrointestinal infection by certain bacteria (e.g., Salmonella, Shigella, Yersinia, Campylobacter and certain forms of Escherichia coli) can be complicated by a form of arthritis which can also produce spinal fusion that is not the result of injury or direct infection of the vertebrae (Resnick 2002; Rothschild and Martin 2006). Could Plioplatecarpus and Clidastes have had a lifestyle that put them in greater contact with such bacteria through water contamination or scavenging contaminated prey?
Pathologies preserved in fossil remains provide valuable insight into interactions between various species and the conditions in which they lived. In mosasaurs, fusion of caudal vertebrae as the result of a non-fatal bite by a shark or other predator, provides evidence of infections that occurred during the healing process, as well as the fact that the mosasaurs lived long enough for the wound to heal.
It is clear that the variety of bridging (marginal or non-marginal was independent of derivation and cannot be utilized to distinguish between post-traumatic/infectious fusion and that related to spondyloarthropathy. It is the presence of associated phenomenon (disorganized architecture, filigree response and sinus drainage) that allows recognition of infection. Traumatic (usually infectious) fusion transcended phylogeny (Plioplatecarpus and Ectenosaurus, excepted), independent of size and habitat. Most of the occurrences of fused vertebrae appear to be attributable to shark attacks. Large and relatively slow-moving mosasaurs seem to have been a popular target for Cretaceous sharks. FHSM VP-13750 certainly exemplifies this propensity.
Pathologies preserved in fossil remains provide valuable insight into interactions between various species and the conditions in which they lived. In mosasaurs, fusion of caudal vertebrae as the result of a non-fatal bite by a shark or other predator, provides evidence of infections that occurred during the healing process, as well as the fact that the mosasaurs lived long enough for the wound to heal.
It is clear that the variety of bridging (marginal or non-marginal was independent of derivation and cannot be utilized to distinguish between post-traumatic/infectious fusion and that related to spondyloarthropathy. It is the presence of associated phenomenon (disorganized architecture, filigree response and sinus drainage) that allows recognition of infection. Traumatic (usually infectious) fusion transcended phylogeny (Plioplatecarpus and Ectenosaurus, excepted), independent of size and habitat. Most of the occurrences of fused vertebrae appear to be attributable to shark attacks. Large and relatively slow-moving mosasaurs seem to have been a popular target for Cretaceous sharks. FHSM VP-13750 certainly exemplifies this propensity.
-----
4. Calcium isotopic analysis - a case study for the extinction of Mosasaurs
Publication for reference: Calcium Isotopic Evidence for Vulnerable Marine Ecosystem Structure Prior to the K/Pg Extinction
SUMMARY
The collapse of marine ecosystems during the end-Cretaceous mass extinction involved the base of the food chain [1] up to ubiquitous vertebrate apex predators [2, 3, 4, 5]. Large marine reptiles became suddenly extinct at the Cretaceous-Paleogene (K/Pg) boundary, whereas other contemporaneous groups such as bothremydid turtles or dyrosaurid crocodylomorphs, although affected at the familial, genus, or species level, survived into post-crisis environments of the Paleocene [5, 6, 7, 8, 9] and could have found refuge in freshwater habitats [10, 11, 12]. A recent hypothesis proposes that the extinction of plesiosaurians and mosasaurids could have been caused by an important drop in sea level [13]. Mosasaurids are unusually diverse and locally abundant in the Maastrichtian phosphatic deposits of Morocco, and with large sharks and one species of elasmosaurid plesiosaurian recognized so far, contribute to an overabundance of apex predators [3, 7, 14, 15]. For this reason, high local diversity of marine reptiles exhibiting different body masses and a wealth of tooth morphologies hints at complex trophic interactions within this latest Cretaceous marine ecosystem. Using calcium isotopes, we investigated the trophic structure of this extinct assemblage. Our results are consistent with a calcium isotope pattern observed in modern marine ecosystems and show that plesiosaurians and mosasaurids indiscriminately fall in the tertiary piscivore group. This suggests that marine reptile apex predators relied onto a single dietary calcium source, compatible with the vulnerable wasp-waist food webs of the modern world [16]. This inferred peculiar ecosystem structure may help explain plesiosaurian and mosasaurid extinction following the end-Cretaceous biological crisis.
Citation: Martin, J. E., Vincent, P., Tacail, T., Khaldoune, F., Jourani, E., Bardet, N., & Balter, V. (2017). Calcium isotopic evidence for vulnerable marine ecosystem structure prior to the K/Pg extinction. Current Biology, 27(11), 1641-1644.
Full read: www.sciencedirect.com/science/article/pii/S0960982217304876#bib16
-----
Findings:
In the studied fossil assemblage (Figure 1), the least fractionated δ44/42Ca value is represented by the marine turtle group (–0.40 ± 0.04‰ 1SD, n = 5) and shows a Δ44/42Ca relative to Late Maastrichtian seawater of −0.70‰, which is identical to the Δ44/42Ca between modern turtles and modern seawater. A second fossil group includes pycnodonts and teleost fishes with the barracuda-like Enchodus as well as two sawsharks of the genus Pristiphorus and has an intermediate mean value (–0.57 ± 0.12‰ 1SD, n = 8) and a Δ44/42Ca relative to Late Maastrichtian seawater of –0.87‰, which is indistinguishable from the Δ44/42Ca value observed between the modern primary piscivore group and modern seawater (–0.87‰). The group containing the large sharks Cretolamna, Squalicorax, and Carcharias (–0.86 ± 0.17‰ 1SD, n = 7) has a Δ44/42Ca relative to Late Maastrichtian seawater of –1.16‰ and is thus very close to the Δ44/42Ca between modern tertiary piscivores and modern seawater (–1.13‰). Marine reptile calcium isotope values strongly overlap with the large sharks but display slightly lower values on average (–0.93 ± 0.12‰ 1SD, n = 31). Values for the elasmosaurid plesiosaurian and the five mosasaurid genera are indistinguishable, suggesting that they indiscriminately fed on the same calcium source. - Martin et al (2017)
Figure 1. δ44/42Ca Variability in Marine Ecosystems of the Modern World and the Maastrichtian
δ44/42Ca data are given in ‰ relative to ICP Ca Lyon standard. For the modern dataset (A), data compilation is from the literature (see [19] for details), with the exception of modern marine turtles (this study). The fossil Maastrichtian dataset (B) is entirely new to this study. 1, modern marine turtles; 2, zooplanktivores; 3, primary piscivores; 4, secondary piscivores; 5, tertiary piscivores; 7, Maastrichtian marine turtles; 8, Maastrichtian primary piscivores; 9, Maastrichtian tertiary piscivores; 10, mosasaurs; 11, plesiosaurs. Triangles represent invertebrates; circles represent marine reptiles; squares represent teleost fishes; diamonds represent elasmobranchs. Error bars represent 2SD. See also Table S1.
-----
Extinction hypothesis:
Here, the structure of the Late Maastrichtian marine ecosystem, as inferred from calcium isotopes, differs in its composition from modern assemblages where apex predators are represented by birds and mammals but also by selachians. By contrast, our data point to a single dietary source for apex predators, suggesting a bottom-up control, with a considerable biomass sustaining an abundant and diversified assemblage of large apex predators. A comparable structure, known as a “wasp waist,” is observed in modern upwelling environments, where a large biomass of poorly diversified forage organisms (the intermediate trophic level) exerts a bottom-up control on diverse apex predators and exerts a top-down control on invertebrate preys [16, 25]. This inference is in perfect agreement with the sedimentological data from the study sites [26], which indicate that the studied fossiliferous phosphate strata were deposited under an upwelling environment. We therefore interpret our results as providing strong evidence for a wasp-waist structure in the studied Late Maastrichtian assemblage. As a caveat, it should be stressed that the structure of modern upwelling ecosystems has not yet been investigated using calcium isotopes.
Our results have implications for the extinction of marine reptiles at the K/Pg boundary. The latest Cretaceous was marked by a major expansion of phosphate deposits in the intertropical zone, interpreted as reflecting intensified upwellings related to the opening of the Atlantic Ocean [26]. In parallel, plesiosaurians and mosasaurids became particularly widespread, diversified, and abundant. Although the wasp-waist structure evidenced by our data for the Moroccan upwelling ecosystem deserves further testing in coeval marine assemblages, it can be reasonably hypothesized that a similar structure characterized many if not all of these low-latitude upwelling ecosystems. If correct, this widespread wasp-waist structure may have rendered the latest Cretaceous ecosystems particularly vulnerable to environmental perturbations of the K/Pg. As a modern example, the drop in abundance and diversity of apex predators in modern upwelling environments is a result of sardine and anchovy overexploitation by industrial fisheries of the late 19th century [16]. Accordingly, the collapse of phytoplankton at the K/Pg boundary [1, 27] likely had a severe impact on intermediate and higher trophic levels occupied by selachians and marine reptiles [6]. This collapse would have been particularly detrimental to plesiosaurians and mosasaurids, which possessed a higher metabolic rate than crocodylomorphs or selachians [28], necessitating substantial caloric intake, hence a corresponding biomass available as prey. This may have been especially sustainable in upwelling zones, which are highly productive environments but are extremely vulnerable ecosystems in contrast to reef ecosystems [29]. The sudden environmental catastrophe disrupting primary productivity at the K/Pg boundary, combined with a widespread and vulnerable type of ecosystem, likely had drastic effects across apex predator communities. Together with models inferring biotic interactions in Cretaceous terrestrial ecosystems [30], our results highlight that the collapse of Maastrichtian marine ecosystems was timely and could have been partly explained by their vulnerable nature, characterized by a wasp-waist structure. - Martin et al (2017)
Our results have implications for the extinction of marine reptiles at the K/Pg boundary. The latest Cretaceous was marked by a major expansion of phosphate deposits in the intertropical zone, interpreted as reflecting intensified upwellings related to the opening of the Atlantic Ocean [26]. In parallel, plesiosaurians and mosasaurids became particularly widespread, diversified, and abundant. Although the wasp-waist structure evidenced by our data for the Moroccan upwelling ecosystem deserves further testing in coeval marine assemblages, it can be reasonably hypothesized that a similar structure characterized many if not all of these low-latitude upwelling ecosystems. If correct, this widespread wasp-waist structure may have rendered the latest Cretaceous ecosystems particularly vulnerable to environmental perturbations of the K/Pg. As a modern example, the drop in abundance and diversity of apex predators in modern upwelling environments is a result of sardine and anchovy overexploitation by industrial fisheries of the late 19th century [16]. Accordingly, the collapse of phytoplankton at the K/Pg boundary [1, 27] likely had a severe impact on intermediate and higher trophic levels occupied by selachians and marine reptiles [6]. This collapse would have been particularly detrimental to plesiosaurians and mosasaurids, which possessed a higher metabolic rate than crocodylomorphs or selachians [28], necessitating substantial caloric intake, hence a corresponding biomass available as prey. This may have been especially sustainable in upwelling zones, which are highly productive environments but are extremely vulnerable ecosystems in contrast to reef ecosystems [29]. The sudden environmental catastrophe disrupting primary productivity at the K/Pg boundary, combined with a widespread and vulnerable type of ecosystem, likely had drastic effects across apex predator communities. Together with models inferring biotic interactions in Cretaceous terrestrial ecosystems [30], our results highlight that the collapse of Maastrichtian marine ecosystems was timely and could have been partly explained by their vulnerable nature, characterized by a wasp-waist structure. - Martin et al (2017)
-----