


Summary of osteophagy in carnivorous reptiles
Feb 11, 2022 4:57:34 GMT 5
Life, theropod, and 1 more like this
Post by Infinity Blade on Feb 11, 2022 4:57:34 GMT 5
Osteophagy (consumption of bones) is a well known behavior among a number of carnivorous mammals, extant and extinct (including, but not limited to, hyaenids, wolverines, Tasmanian devil, borophagines, entelodonts, certain mesonychids). Such predators consistently share certain cranial and dental traits in order to efficiently consume bone, such as blunt premolars for cracking bones, strong jaw muscles, etc.
By contrast, most carnivorous reptiles today (including birds) don’t seem to be in the habit of regularly consuming bone. It’s been noted before that they tend not to have the specialized heterodont dentition mammals have, including hammer-like bone-cracking premolars. Additionally, “extant carnivorous reptiles (Sauria—including birds [Neornithes]) typically possess non-occluding teeth, or in the case of modern birds lack them entirely, and cannot generate sufficient stress distributions to fragment bones. Instead, they consume small carcasses in their entirety and large skeletal elements through dismemberment.” (Gignac & Erickson, 2017). Does this mean bone-smashing and eating carnivorous reptiles aren’t really much of a thing?
Not at all. In addition to the one highly specialized bone-eating bird alive today (the lammergeier), the fossil record reveals multiple lineages of osteophagous carnivorous reptiles. And interestingly, they didn’t all converge on the same solution. This thread is simply a summary of known examples of osteophagous carnivorous reptiles, and the different methods they use(d) to practice the behavior.
Tyrannosauridae:
These are probably the most well-known examples. As we’re all very much aware, tyrannosaurids not only bit extremely hard, but they also exerted their bite force on pointed tooth tips, generating high pressures on them (Gignac & Erickson, 2017*).
*Keep in mind that much higher bite force estimates for Tyrannosaurus exist, some of which came after this study (Bates & Falkingham, 2012; Snively et al., 2015; Cost et al., 2019; Rowe & Snively, 2021).
After penetration, the incrassate, broadly-expanded tooth crowns created cracks. In addition, the front and back carinae present on tyrannosaurid teeth also propagated cracks in the bone in the direction they faced (see the video below to see what this looks like); these cracks would be directed towards the cracks formed by the adjacent tooth/carina. That is to say, the front carina of one tooth would generate a crack running forwards, and the back carina of the tooth in front would generate a crack running backwards. Tyrannosaurids had an entire battery of these teeth, generating a long cracking arcade that would split bones apart (Gignac & Erickson, 2017).
www.bbc.co.uk/programmes/p05rqcl5
Also, “the dental and palatal anatomy also provided for three- and four-point loading configurations that facilitated localized and whole-element bone shear” (Gignac & Erickson, 2017).
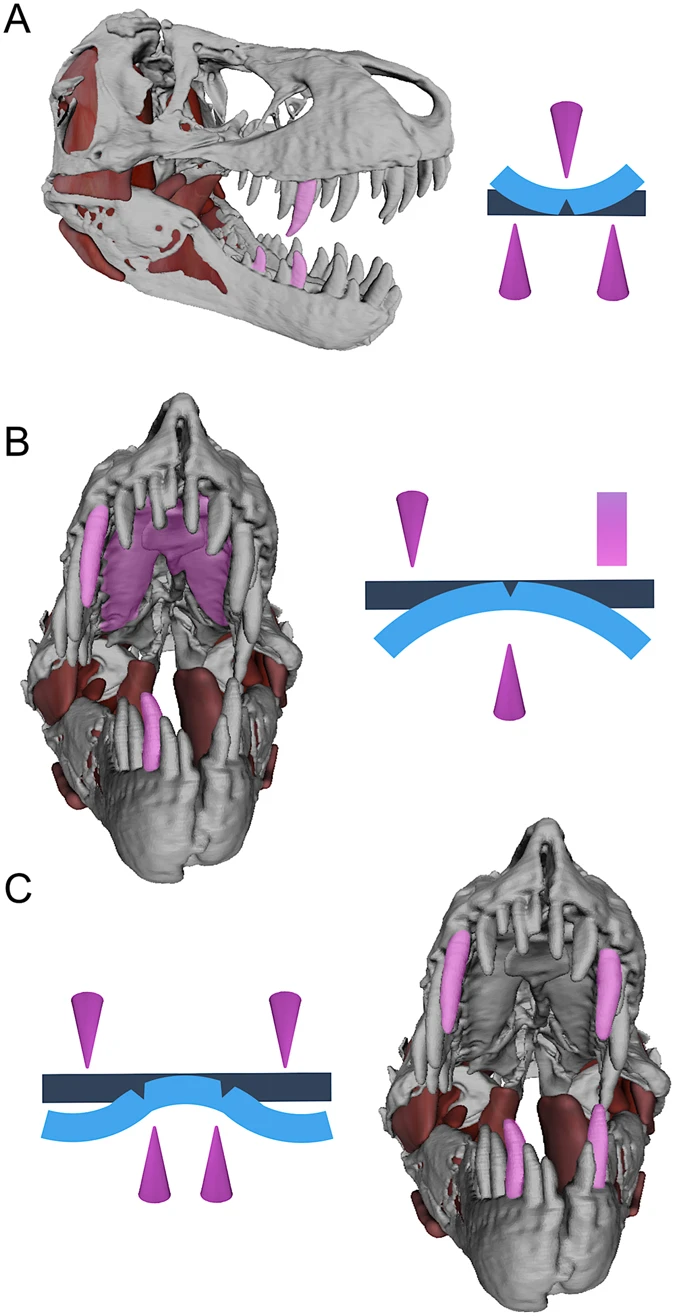
With this jaw and dental anatomy, tyrannosaurids were easily capable of crushing and processing large dinosaur bones (Erickson & Olson, 1996). One coprolite attributed to Tyrannosaurus rex was 30-50% bone by volume (Chin et al., 1998).
Abelisauridae (at least Majungasaurus crenatissimus):
Majungasaurus crenatissimus, and perhaps other abelisaurids, took a very different approach to osteophagy than the tyrannosaurids. It lacked the incrassate teeth of tyrannosaurids (although, it did have short-crowned teeth), but it had a very quick tooth replacement rate, estimated to be 56 days. This is significantly quicker than in Allosaurus and Ceratosaurus (estimated to be over 100 days), and far quicker than in tyrannosaurids such a T. rex (estimated to be 777 days). This is believed to be an adaptation to osteophagy, as evidenced by extensive tooth markings on both other Majungasaurus bones or the bones of sympatric herbivores. Other abelisaurids may have been similar to Majungasaurus, as some specimens have been found to have been packed with unerupted teeth in their maxillae (D'Emic et al., 2019).
Smok wawelski:
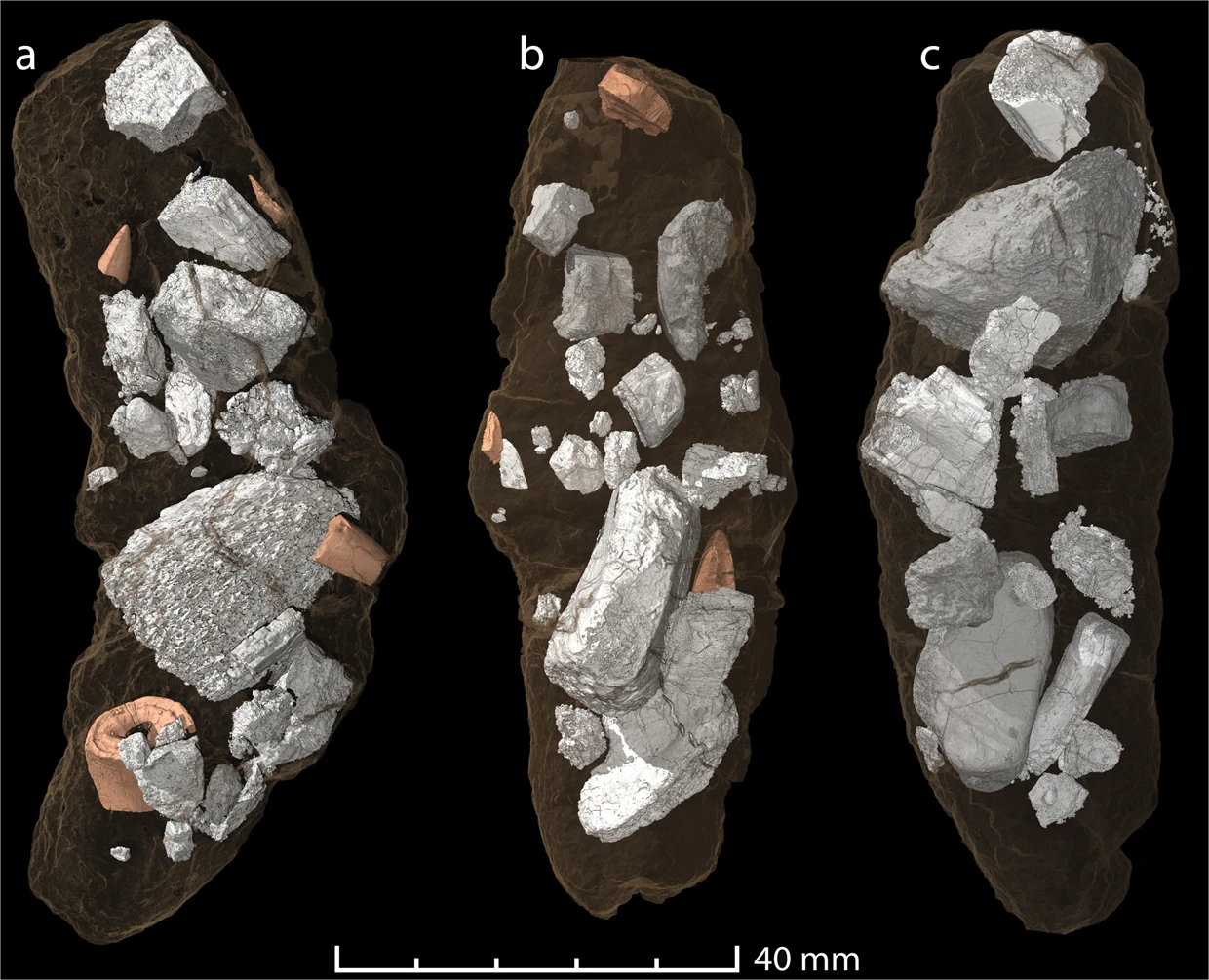
Smok is a rather poorly understood genus of predatory archosaur that lived in the Triassic of Poland. We currently lack sufficient remains to conclusively tell whether it is a dinosaur or pseudosuchian. Nevertheless, teeth are known from this animal. The teeth were recurved and serrated, characteristic of predatory archosaurs (Niedźwiedzki et al., 2011). More recently, coprolites attributable to Smok (based on size, contents, and even fragments of matching teeth) have been found, being 50% bone by volume. Bones from a variety of prey items were found from these coprolites, including those of dicynodonts, suggesting tyrannosaurid-like osteophagy. (Qvarnström et al., 2019).
Razanandrongobe sakalavae:
This species is a gigantic mesoeucrocodylian that lived in the Middle Jurassic of Madagascar, and the earliest known notosuchian. When it was first discovered its phylogenetic affinities were less clear, but some material, like the dentition and maxilla, was known. Even when it was first discovered it was strongly suspected this predator crushed bones given the structure of its teeth, maxilla, and heavy wear on the teeth (Maganuco et al., 2006).

The massive, robust jaw bones that were later discovered support this idea (Dal Sasso et al., 2017). Additionally, the mesial incisiform teeth of Razanandrongobe look like the premaxillary teeth of tyrannosaurids. Denticle shape and size also resemble that seen in tyrannosaurids (Dal Sasso et al., 2017).

Snout reconstruction from Dal Sasso et al. (2017).
Allosauridae:
Any carnosaur would seem like an odd entry into this list, as these animals had laterally compressed teeth and less emphasis on jaw adductor strength than say, tyrannosaurids. Nor do allosaurids possess the extreme tooth replacement rates seen in abelisaurids like Majungasaurus. So why include allosaurids? Well, this is why.
(Luckily Duane Nash copied and pasted all the relevant info of this abstract on his blog->, as the direct link to this abstract now leads to a 404 page)
So what we have is a massive Allosaurus coprolite that is ~50% bone by volume. This is just as much as the Smok coprolite, and at least as much, if not more, than the T. rex coprolite. Also, the bone fragments were processed into smaller portions, not swallowed whole like carnivorous reptiles usually do. If that doesn’t qualify Allosaurus as an osteophage, I don’t know what will.
Allosaurus had the ability to remove large sections of bone with its jaws and teeth, as evidenced by a Camarasaurus ilium with a section removed by an Allosaurus’ teeth (Chure et al., 1998).

It certainly had the ability to rake bone with its teeth and produce furrows (link->). However, the mesial teeth of Allosaurus were also mesiodistally thick, allowing them to endure high stresses and to engage in bone crunching (Hendrickx et al., 2020).
Gypaetus barbatus:
Of course, there is one carnivorous sauropsid (a bird, in this case) alive today that is, in fact, a specialized osteophage. The lammergeier (or bearded vulture) is the only vertebrate (source->) whose diet consists almost exclusively of bones, forming up to 85% of its diet (Ferguson-Lees & Christie, 2001). Smaller bones are swallowed whole (as is usual for recent carnivorous archosaurs; Qvarnström et al. 2019) or bitten through if brittle enough. Larger bones are carried up into the air with the feet and dropped from heights of 50-80 meters onto hard, flat rocks, as many times as necessary (Ferguson-Lees & Christie, 2001).

Image source->
By contrast, most carnivorous reptiles today (including birds) don’t seem to be in the habit of regularly consuming bone. It’s been noted before that they tend not to have the specialized heterodont dentition mammals have, including hammer-like bone-cracking premolars. Additionally, “extant carnivorous reptiles (Sauria—including birds [Neornithes]) typically possess non-occluding teeth, or in the case of modern birds lack them entirely, and cannot generate sufficient stress distributions to fragment bones. Instead, they consume small carcasses in their entirety and large skeletal elements through dismemberment.” (Gignac & Erickson, 2017). Does this mean bone-smashing and eating carnivorous reptiles aren’t really much of a thing?
Not at all. In addition to the one highly specialized bone-eating bird alive today (the lammergeier), the fossil record reveals multiple lineages of osteophagous carnivorous reptiles. And interestingly, they didn’t all converge on the same solution. This thread is simply a summary of known examples of osteophagous carnivorous reptiles, and the different methods they use(d) to practice the behavior.
Tyrannosauridae:
These are probably the most well-known examples. As we’re all very much aware, tyrannosaurids not only bit extremely hard, but they also exerted their bite force on pointed tooth tips, generating high pressures on them (Gignac & Erickson, 2017*).
*Keep in mind that much higher bite force estimates for Tyrannosaurus exist, some of which came after this study (Bates & Falkingham, 2012; Snively et al., 2015; Cost et al., 2019; Rowe & Snively, 2021).
After penetration, the incrassate, broadly-expanded tooth crowns created cracks. In addition, the front and back carinae present on tyrannosaurid teeth also propagated cracks in the bone in the direction they faced (see the video below to see what this looks like); these cracks would be directed towards the cracks formed by the adjacent tooth/carina. That is to say, the front carina of one tooth would generate a crack running forwards, and the back carina of the tooth in front would generate a crack running backwards. Tyrannosaurids had an entire battery of these teeth, generating a long cracking arcade that would split bones apart (Gignac & Erickson, 2017).
www.bbc.co.uk/programmes/p05rqcl5
Also, “the dental and palatal anatomy also provided for three- and four-point loading configurations that facilitated localized and whole-element bone shear” (Gignac & Erickson, 2017).
With this jaw and dental anatomy, tyrannosaurids were easily capable of crushing and processing large dinosaur bones (Erickson & Olson, 1996). One coprolite attributed to Tyrannosaurus rex was 30-50% bone by volume (Chin et al., 1998).
Abelisauridae (at least Majungasaurus crenatissimus):
Majungasaurus crenatissimus, and perhaps other abelisaurids, took a very different approach to osteophagy than the tyrannosaurids. It lacked the incrassate teeth of tyrannosaurids (although, it did have short-crowned teeth), but it had a very quick tooth replacement rate, estimated to be 56 days. This is significantly quicker than in Allosaurus and Ceratosaurus (estimated to be over 100 days), and far quicker than in tyrannosaurids such a T. rex (estimated to be 777 days). This is believed to be an adaptation to osteophagy, as evidenced by extensive tooth markings on both other Majungasaurus bones or the bones of sympatric herbivores. Other abelisaurids may have been similar to Majungasaurus, as some specimens have been found to have been packed with unerupted teeth in their maxillae (D'Emic et al., 2019).
Smok wawelski:
Smok is a rather poorly understood genus of predatory archosaur that lived in the Triassic of Poland. We currently lack sufficient remains to conclusively tell whether it is a dinosaur or pseudosuchian. Nevertheless, teeth are known from this animal. The teeth were recurved and serrated, characteristic of predatory archosaurs (Niedźwiedzki et al., 2011). More recently, coprolites attributable to Smok (based on size, contents, and even fragments of matching teeth) have been found, being 50% bone by volume. Bones from a variety of prey items were found from these coprolites, including those of dicynodonts, suggesting tyrannosaurid-like osteophagy. (Qvarnström et al., 2019).
Razanandrongobe sakalavae:
This species is a gigantic mesoeucrocodylian that lived in the Middle Jurassic of Madagascar, and the earliest known notosuchian. When it was first discovered its phylogenetic affinities were less clear, but some material, like the dentition and maxilla, was known. Even when it was first discovered it was strongly suspected this predator crushed bones given the structure of its teeth, maxilla, and heavy wear on the teeth (Maganuco et al., 2006).
The massive, robust jaw bones that were later discovered support this idea (Dal Sasso et al., 2017). Additionally, the mesial incisiform teeth of Razanandrongobe look like the premaxillary teeth of tyrannosaurids. Denticle shape and size also resemble that seen in tyrannosaurids (Dal Sasso et al., 2017).
Snout reconstruction from Dal Sasso et al. (2017).
Allosauridae:
Any carnosaur would seem like an odd entry into this list, as these animals had laterally compressed teeth and less emphasis on jaw adductor strength than say, tyrannosaurids. Nor do allosaurids possess the extreme tooth replacement rates seen in abelisaurids like Majungasaurus. So why include allosaurids? Well, this is why.
A LARGE MEAT-EATING DINOSAUR COPROLITE FROM THE JURASSIC MORRISON FORMATION OF UTAH
Author(s): STONE, Dwayne D., Dept. of Geology, Marietta College, Marietta, OH 45750; CRISP, Edward L., Geology Dept., West Virginia University at Parkersburg, Parkersburg, WV 26101, ecrisp@alpha.wvup.wvnet.edu; BISHOP, John R., Rt. 2 Box 137, Ravenswood, WV 26164
Keywords: Coprolite, Jurassic, Morrison Formation, Allosaurus
A theropod dinosaur coprolite has been excavated in Emery County, Utah from a red-brown mudstone of the Upper Jurassic Brushy Basin Member of the Morrison Formation. The coprolite is nearly complete and is divisible into two parts, the main mass and a dribbling zone. The well-indurated main mass, which tapers at both ends, is 1.52 m in length, 0.457 m in maximum width, and 10.2 cm thick. The 1.52 m long dribbling zone consists of small isolated coprolites that curve towards the end of the dribbles. The coprolite geometry is interpreted to indicate that the main mass was defecated first, then the animal walked forward to release smaller amounts of feces. The coprolite consists of dark gray bone fragments, ranging from pebble to sand size, in a red-brown matrix. Bone fragments represent about 50% of the mass and have broken, jagged ends, possibly indicating breakage by the biting action of a carnivore. XRD analysis of the bone fragments and matrix reveal that both are primarily composed of carbonate fluorapatite. The large size of the coprolite and its geometry and stratigraphic location indicate that it represents fecal droppings from a large Allosaurus. Further support for this hypothesis is the fact that a broken distal end of an Allosaurus tooth was found within the coprolite, indicating that during mastication a tooth was broken and ingested. The carnivore did not grab a portion of flesh and bone from a prey animal and then swallow it whole. Instead the eater masticated its meal and broke the bones into smaller portions. This is the largest and oldest theropod dinosaur coprolite known. Work is continuing on a second theropod coprolite higher in the section at this location and pieces of theropod coprolites from two additional localities have been identified. Morrison Formation theropod coprolites are no longer considered to be absent or scarce and future searching should reveal additional large specimens.
Author(s): STONE, Dwayne D., Dept. of Geology, Marietta College, Marietta, OH 45750; CRISP, Edward L., Geology Dept., West Virginia University at Parkersburg, Parkersburg, WV 26101, ecrisp@alpha.wvup.wvnet.edu; BISHOP, John R., Rt. 2 Box 137, Ravenswood, WV 26164
Keywords: Coprolite, Jurassic, Morrison Formation, Allosaurus
A theropod dinosaur coprolite has been excavated in Emery County, Utah from a red-brown mudstone of the Upper Jurassic Brushy Basin Member of the Morrison Formation. The coprolite is nearly complete and is divisible into two parts, the main mass and a dribbling zone. The well-indurated main mass, which tapers at both ends, is 1.52 m in length, 0.457 m in maximum width, and 10.2 cm thick. The 1.52 m long dribbling zone consists of small isolated coprolites that curve towards the end of the dribbles. The coprolite geometry is interpreted to indicate that the main mass was defecated first, then the animal walked forward to release smaller amounts of feces. The coprolite consists of dark gray bone fragments, ranging from pebble to sand size, in a red-brown matrix. Bone fragments represent about 50% of the mass and have broken, jagged ends, possibly indicating breakage by the biting action of a carnivore. XRD analysis of the bone fragments and matrix reveal that both are primarily composed of carbonate fluorapatite. The large size of the coprolite and its geometry and stratigraphic location indicate that it represents fecal droppings from a large Allosaurus. Further support for this hypothesis is the fact that a broken distal end of an Allosaurus tooth was found within the coprolite, indicating that during mastication a tooth was broken and ingested. The carnivore did not grab a portion of flesh and bone from a prey animal and then swallow it whole. Instead the eater masticated its meal and broke the bones into smaller portions. This is the largest and oldest theropod dinosaur coprolite known. Work is continuing on a second theropod coprolite higher in the section at this location and pieces of theropod coprolites from two additional localities have been identified. Morrison Formation theropod coprolites are no longer considered to be absent or scarce and future searching should reveal additional large specimens.
(Luckily Duane Nash copied and pasted all the relevant info of this abstract on his blog->, as the direct link to this abstract now leads to a 404 page)
So what we have is a massive Allosaurus coprolite that is ~50% bone by volume. This is just as much as the Smok coprolite, and at least as much, if not more, than the T. rex coprolite. Also, the bone fragments were processed into smaller portions, not swallowed whole like carnivorous reptiles usually do. If that doesn’t qualify Allosaurus as an osteophage, I don’t know what will.
Allosaurus had the ability to remove large sections of bone with its jaws and teeth, as evidenced by a Camarasaurus ilium with a section removed by an Allosaurus’ teeth (Chure et al., 1998).
It certainly had the ability to rake bone with its teeth and produce furrows (link->). However, the mesial teeth of Allosaurus were also mesiodistally thick, allowing them to endure high stresses and to engage in bone crunching (Hendrickx et al., 2020).
If the dentition of Sinraptor dongi is obviously adapted for a carnivorous diet, several features suggest that its teeth were suited for a predatory lifestyle. Similar to those of abelisaurids and tyrannosaurids, the mesial teeth of Sinraptor dongi and Allosaurus are mesiodistally thick and were able to endure high mechanical stresses (Hendrickx et al. 2019). Robust mesial dentition appears to be adapted towards diets involving bone crunching and bone biting, with high degrees of torsion applied on mesial teeth (Reichel 2010, 2012). Such a mesial dentition was suited for gripping and pulling on prey in a bite-and-hold manner (Sampson and Witmer 2007; Reichel 2012). If mesial teeth with lingually deflected carinae and D- and U-shaped cross sections are perfectly suited for defleshing carcasses (Reichel 2012), the wide distances between mesial and distal carinae in mesial teeth, associated with spiralling mesial carinae and deflected distal carinae in anterior lateral teeth, would make wide cuts, keeping the wounds open and ultimately causing fatal injuries to the prey (Bakker 1998; Reichel 2012; Hendrickx et al. 2019). Additional evidence supports Sinraptor dongi as a predator. The large number of partially healed bite wounds seen on the cranial material of Sinraptor dongi (i.e., 28 tooth strike lesions distributed on the maxilla, jugal, dentary, prearticular, and possibly the surangular; Tanke and Currie 1998; Fig. 2B) was interpreted by Tanke and Currie (1998) to be indicative of aggressive inter- or intraspecific biting. Because Sinraptor dongi appears to be the only large-bodied theropod from the upper beds of the Shishugou Formation, intraspecific head- and face-biting behaviour most likely occurred in this taxon. If true, this supports the fact that the dentition of Sinraptor dongi was capable of enduring tooth-to-bone contact and was consequently well suited to a predatory lifestyle. Indeed, extensive spalled surfaces on the crown apices, which reflect flaking of enamel caused by the contact between the crown and food (Schubert and Ungar 2005), combined with fully worn apices in some mesial and lateral teeth, clearly indicate particularly high forces produced during contact with what was most likely the bones of prey items. Sinraptor dongi, together with Allosaurus, have been hypothesized to be “strike-and-tear/pull feeders”, rapidly striking downwards at smaller prey using the mesial dentition (Snively and Russell 2007). Neck muscle morphologies in Sinraptor dongi and Allosaurus correlate with powerful ventroflexise kinematics, which would facilitate cutting flesh with the upper dentition (Snively and Russell 2007). Allosaurus, whose dentition is very similar to that of Sinraptor dongi, is widely regarded as a predator on the basis of the discovery of a Stegosaurus cervical plate with a bite pattern matching that of Allosaurus, and an Allosaurus caudal vertebra showing a partially healed wound likely caused by a Stegosaurus tail spike (Carpenter et al. 2005). Consequently, Sinraptor dongi is here interpreted as a predator, likely having preyed on medium-sized herbivorous dinosaurs such as the stegosaurid Jiangjunosaurus junggarensis, using its dentition to inflict fatal injuries in a manner similar to that of Allosaurus.
Gypaetus barbatus:
Of course, there is one carnivorous sauropsid (a bird, in this case) alive today that is, in fact, a specialized osteophage. The lammergeier (or bearded vulture) is the only vertebrate (source->) whose diet consists almost exclusively of bones, forming up to 85% of its diet (Ferguson-Lees & Christie, 2001). Smaller bones are swallowed whole (as is usual for recent carnivorous archosaurs; Qvarnström et al. 2019) or bitten through if brittle enough. Larger bones are carried up into the air with the feet and dropped from heights of 50-80 meters onto hard, flat rocks, as many times as necessary (Ferguson-Lees & Christie, 2001).
Image source->
