Post by elosha11 on Feb 21, 2013 20:27:50 GMT 5
Spinosaurus aegyptiacus

Life reconstruction of S. aegyptiacus, accounting for all known evidence as of January 2021. © @ Chris Masna
Temporal range: Cretaceous; Cenomanian (99-93.5 million years ago)
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Cephalochordata
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Clade: Euteleostomi
Clade: Sarcopterygii
Clade: Rhipidistia
Clade: Tetrapodomorpha
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Class: Reptilia or Clade: Sauropsida
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Order: Saurischia
Clade: Eusaurischia
Suborder: Theropoda
Clade: Neotheropoda
Clade: Averostra
Clade: Tetanurae
Clade: Orionides
Clade: Avetheropoda
Superfamily: †Megalosauroidea
Clade: †Megalosauria
Family: †Spinosauridae
Genus: †Spinosaurus
Species: †S. aegyptiacus
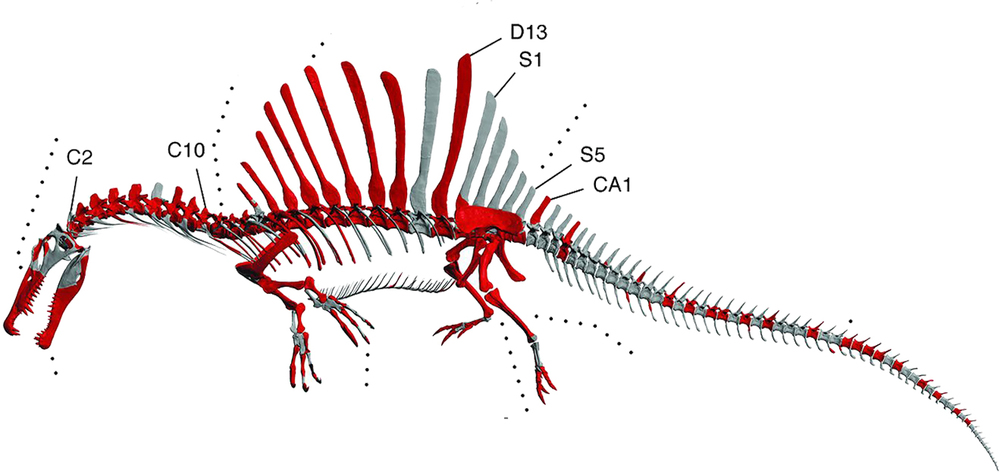
Spinosaurus is an extinct genus of theropod dinosaur that lived in North Africa from 99-93.5 million years ago.
Locality:
Spinosaurus remains (or remains comparable to it) have been uncovered in Algeria, Tunisia, Egypt, Morocco, and Niger.[1]
Taxonomy:
Two Spinosaurus species have been named based on the region where they were discovered: Spinosaurus aegyptiacus (Egyptian spine lizard) and Spinosaurus maroccanus (Moroccan spine lizard). The latter is nowadays regarded as a junior synonym of Sigilmassasaurus brevicollis.[2]
Description and paleobiology:
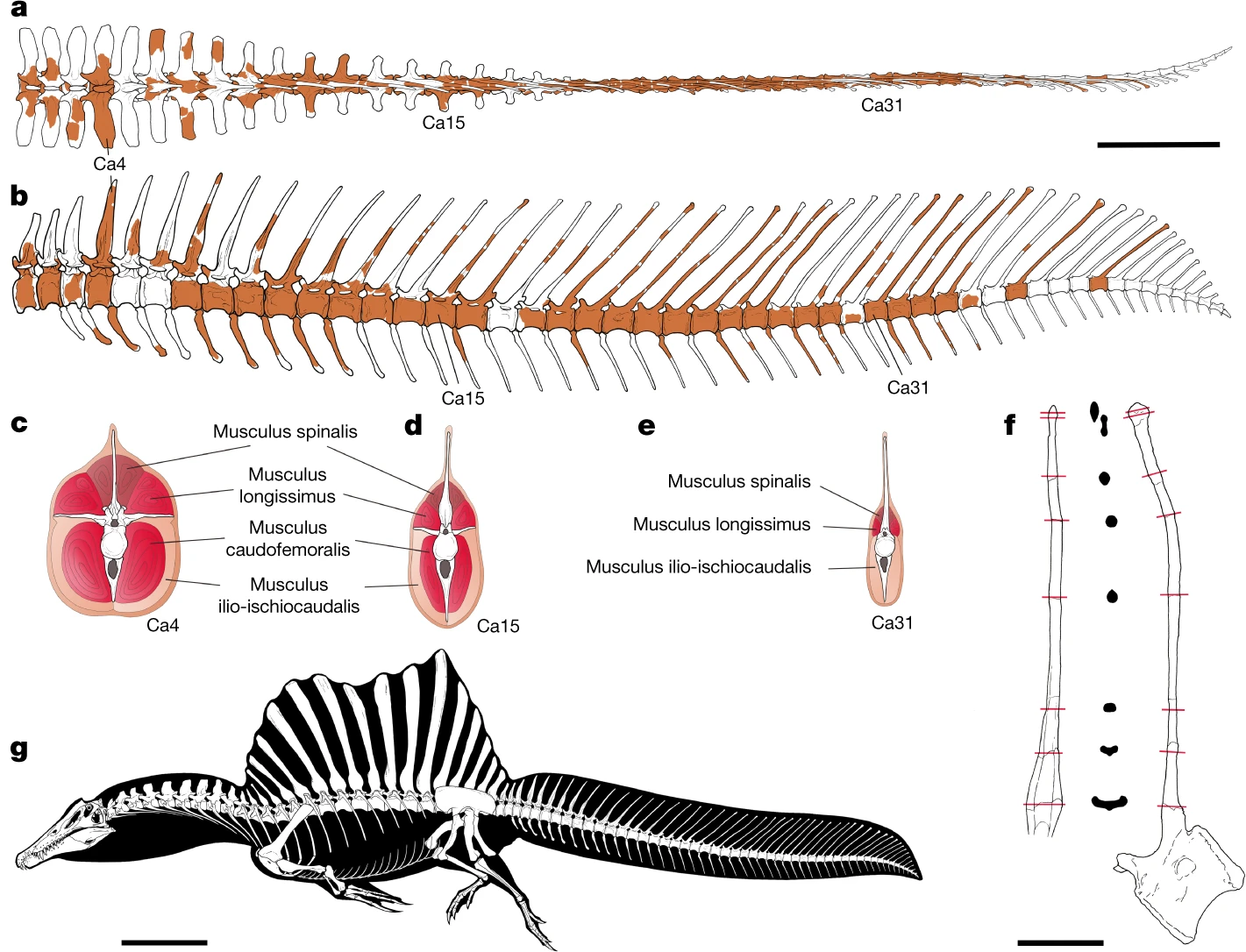
Spinosaurus was named for the elongated neural spines that protruded from its back, forming a sail. One recent hypothesis proposes that the sail was used to improve maneuverability while swimming underwater, while providing hydrodynamic fulcrum needed to make powerful neck and tail blows (akin to those made by sailfish and thresher sharks) to stun, injure, or kill prey. The sail may also have served to help herd shoals of prey.[3] This hypothesis was rejected by a more recent paper, which claims that the torso and sail were too inflexible to function akin to that of a sailfish (which are flexible, collapsable, and mediolaterally thin).[4]
Spinosaurus exhibits a number of adaptations to a semiaquatic lifestyle[5][6]. There is still debate as to whether Spinosaurus was a shoreline generalist or an aquatic pursuit specialist.[4][5][6]
S. aegyptiacus is one of the largest known theropods, with a body length estimated to reach 15 meters.[5] Specimen FSAC-KK 11888 in particular was estimated to weigh anywhere between 3,219-4,173 kg.[6 supp. info] The non-specific gravity of Spinosaurus was estimated to be similar to that of crocodilians, at 1.05, making it slightly more dense than water.[7]
The cranial anatomy of Spinosaurus has also received attention from paleontologists. The anterior rostrum of Spinosaurus and Baryonyx both absolutely outperform modern crocodilian taxa in their ability to withstand stress, due to their large size. Despite differences in size and morphology, the anterior rostra of both Spinosaurus and Baryonyx perform similarly, suggesting similar feeding methods between the two. When the effects of size are removed, Baryonyx still outperforms all other taxa in dorsoventral bending, but Spinosaurus' performance falls between that of the alligator and gharial. In mediolateral bending, Baryonyx performs intermediately between the alligator and gharial, while Spinosaurus performs worse than all crocodilian taxa (a later study found that the anterior rostrum of Baryonyx performs similarly in bending to that of a Nile crocodile[8] with a total skull length of 50.1 cm[9]). In torsion, Baryonyx performs intermediately between the alligator and all other crocodilians, while Spinosaurus performs equal to or worse than the gharial and the slender-snouted crocodile. It should be noted that only the anterior rostrum was tested, and the results must be considered in this context.[10]
As a result, Spinosaurus and its kin seem to have been more apt to kill smaller prey via dorsoventral shaking. The deeply-rooted teeth and near vertical-sided teeth rows of spinosaurids are ideal for withstanding large dorsoventral bite forces and dissipating the resulting forces throughout the skull. The ability of Spinosaurus to prey on larger, struggling prey was not conferred by a snout specialized in withstanding stress, but by simple size-related advantages.[10]
The mandibular ramus of Spinosaurus was able to move laterally thanks to a mobile mandibular symphysis, which allowed the pharynx to widen. This is similar to jaw mechanics in some pterosaurs and pelicans. Hence, it is thought that Spinosaurus had the ability to swallow large fish and other prey items, with the ability to prey upon pterosaurs and juvenile dinosaurs.[11]
Isotope analysis and taphonomic evidence both support the use of aquatic resources by Spinosaurus.[12][13]

Carved reconstructed of the cranium of Spinosaurus, held by paleoartist Tyler Keillor.
Competition:
Interestingly, large crocodylomorph remains (e.g. Sarcosuchus) dominate Aptian beds in Africa and South America, but are unknown from the Cenomanian of North Africa. The largest crocodylomorph from the latter beds is Elosuchus; with a skull ~100 cm long, it would have been smaller than the giant spinosaurids. Larger crocodylomorphs seem rare in the Kem Kem Beds and Bahariya fauna. This suggests that spinosaurids became freshwater apex predators in North Africa as giant crocodylomorphs declined. Spinosaurids may have either replaced a vacant niche left behind by these crocodylomorphs or actively outcompeted them into extinction.[14]
References:
[1] paleobiodb.org/classic/basicTaxonInfo?taxon_no=53372
[2] Evers SW, Rauhut OWM, Milner AC, McFeeters B, Allain R. 2015. A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco. PeerJ 3:e1323 doi.org/10.7717/peerj.1323
[3] GIMSA, J., SLEIGH, R., & GIMSA, U. (2016). The riddle of Spinosaurus aegyptiacus’ dorsal sail. Geological Magazine, 153(3), 544-547. doi:10.1017/S0016756815000801
[4] Hone, D., & Holtz, T. (2021). Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?. Palaeontologia Electronica.
[5] Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D. M., Zouhri, S., Myhrvold, N., & Iurino, D. A. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science (New York, N.Y.), 345(6204), 1613–1616. doi.org/10.1126/science.1258750
[6] Ibrahim, N., Maganuco, S., Dal Sasso, C. et al. Tail-propelled aquatic locomotion in a theropod dinosaur. Nature 581, 67–70 (2020). doi.org/10.1038/s41586-020-2190-3
[7] Larramendi, A., Paul, G. S., & Hsu, S. Y. (2020). A review and reappraisal of the specific gravities of present and past multicellular organisms, with an emphasis on tetrapods. Anatomical record (Hoboken, N.J. : 2007), 10.1002/ar.24574. Advance online publication. doi.org/10.1002/ar.24574
[8] Foffa, D., Cuff, A. R., Sassoon, J., Rayfield, E. J., Mavrogordato, M. N., & Benton, M. J. (2014). Functional anatomy and feeding biomechanics of a giant Upper Jurassic pliosaur (Reptilia: Sauropterygia) from Weymouth Bay, Dorset, UK. Journal of anatomy, 225(2), 209–219. doi.org/10.1111/joa.12200
[9] Lemanis, R., Jones, A. S., Butler, R. J., Anderson, P. S., & Rayfield, E. J. (2019). Comparative biomechanical analysis demonstrates functional convergence between slender-snouted crocodilians and phytosaurs (No. e27476v1). PeerJ Preprints.
[10] Cuff, A. R., & Rayfield, E. J. (2013). Feeding mechanics in spinosaurid theropods and extant crocodilians. PloS one, 8(5), e65295. doi.org/10.1371/journal.pone.0065295
[11] Hendrickx, C., Mateus, O., & Buffetaut, E. (2016). [url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144695Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa[/url]. PloS one, 11(1), e0144695. doi.org/10.1371/journal.pone.0144695
[12] Amiot, R., Buffetaut, E., Lécuyer, C., Wang, X., Boudad, L., Ding, Z., ... & Zhou, Z. (2010). Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods. Geology, 38(2), 139-142.[13] Beevor, T., Quigley, A., Smith, R. E., Smyth, R. S. H
[13] Ibrahim, N., Zouhri, S., & Martill, D. M. (2021). Taphonomic evidence supports an aquatic lifestyle for Spinosaurus. Cretaceous Research, 117, [104627]. doi.org/10.1016/j.cretres.2020.104627
[14] Arden, T. M., Klein, C. G., Zouhri, S., & Longrich, N. R. (2019). Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurids. Cretaceous Research, 93, 275-284.
Life reconstruction of S. aegyptiacus, accounting for all known evidence as of January 2021. © @ Chris Masna
Temporal range: Cretaceous; Cenomanian (99-93.5 million years ago)
Scientific classification:
Life
Domain: Eukaryota
(unranked): Unikonta
(unranked): Opisthokonta
(unranked): Holozoa
(unranked): Filozoa
Kingdom: Animalia
Subkingdom: Eumetazoa
(unranked): Bilateria
Clade: Nephrozoa
Superphylum: Deuterostomia
Phylum: Chordata
Clade: Cephalochordata
Clade: Craniata
Subphylum: Vertebrata
Infraphylum: Gnathostomata
Clade: Eugnathostomata
Clade: Teleostomi
Clade: Euteleostomi
Clade: Sarcopterygii
Clade: Rhipidistia
Clade: Tetrapodomorpha
Superclass: Tetrapoda
Clade: Reptiliomorpha
Clade: Amniota
Class: Reptilia or Clade: Sauropsida
Clade: Eureptilia
Clade: Romeriida
Clade: Diapsida
Clade: Neodiapsida
Clade: Archelosauria
Clade: Archosauromorpha
Clade: Archosauriformes
Clade: Crurotarsi
Clade: Archosauria
Clade: Avemetatarsalia
Clade: Ornithodira
Clade: Dinosauromorpha
Clade: Dinosauriformes
Clade: Dinosauria
Order: Saurischia
Clade: Eusaurischia
Suborder: Theropoda
Clade: Neotheropoda
Clade: Averostra
Clade: Tetanurae
Clade: Orionides
Clade: Avetheropoda
Superfamily: †Megalosauroidea
Clade: †Megalosauria
Family: †Spinosauridae
Genus: †Spinosaurus
Species: †S. aegyptiacus
Spinosaurus is an extinct genus of theropod dinosaur that lived in North Africa from 99-93.5 million years ago.
Locality:
Spinosaurus remains (or remains comparable to it) have been uncovered in Algeria, Tunisia, Egypt, Morocco, and Niger.[1]
Taxonomy:
Two Spinosaurus species have been named based on the region where they were discovered: Spinosaurus aegyptiacus (Egyptian spine lizard) and Spinosaurus maroccanus (Moroccan spine lizard). The latter is nowadays regarded as a junior synonym of Sigilmassasaurus brevicollis.[2]
Description and paleobiology:
Spinosaurus was named for the elongated neural spines that protruded from its back, forming a sail. One recent hypothesis proposes that the sail was used to improve maneuverability while swimming underwater, while providing hydrodynamic fulcrum needed to make powerful neck and tail blows (akin to those made by sailfish and thresher sharks) to stun, injure, or kill prey. The sail may also have served to help herd shoals of prey.[3] This hypothesis was rejected by a more recent paper, which claims that the torso and sail were too inflexible to function akin to that of a sailfish (which are flexible, collapsable, and mediolaterally thin).[4]
Spinosaurus exhibits a number of adaptations to a semiaquatic lifestyle[5][6]. There is still debate as to whether Spinosaurus was a shoreline generalist or an aquatic pursuit specialist.[4][5][6]
S. aegyptiacus is one of the largest known theropods, with a body length estimated to reach 15 meters.[5] Specimen FSAC-KK 11888 in particular was estimated to weigh anywhere between 3,219-4,173 kg.[6 supp. info] The non-specific gravity of Spinosaurus was estimated to be similar to that of crocodilians, at 1.05, making it slightly more dense than water.[7]
The cranial anatomy of Spinosaurus has also received attention from paleontologists. The anterior rostrum of Spinosaurus and Baryonyx both absolutely outperform modern crocodilian taxa in their ability to withstand stress, due to their large size. Despite differences in size and morphology, the anterior rostra of both Spinosaurus and Baryonyx perform similarly, suggesting similar feeding methods between the two. When the effects of size are removed, Baryonyx still outperforms all other taxa in dorsoventral bending, but Spinosaurus' performance falls between that of the alligator and gharial. In mediolateral bending, Baryonyx performs intermediately between the alligator and gharial, while Spinosaurus performs worse than all crocodilian taxa (a later study found that the anterior rostrum of Baryonyx performs similarly in bending to that of a Nile crocodile[8] with a total skull length of 50.1 cm[9]). In torsion, Baryonyx performs intermediately between the alligator and all other crocodilians, while Spinosaurus performs equal to or worse than the gharial and the slender-snouted crocodile. It should be noted that only the anterior rostrum was tested, and the results must be considered in this context.[10]
As a result, Spinosaurus and its kin seem to have been more apt to kill smaller prey via dorsoventral shaking. The deeply-rooted teeth and near vertical-sided teeth rows of spinosaurids are ideal for withstanding large dorsoventral bite forces and dissipating the resulting forces throughout the skull. The ability of Spinosaurus to prey on larger, struggling prey was not conferred by a snout specialized in withstanding stress, but by simple size-related advantages.[10]
The mandibular ramus of Spinosaurus was able to move laterally thanks to a mobile mandibular symphysis, which allowed the pharynx to widen. This is similar to jaw mechanics in some pterosaurs and pelicans. Hence, it is thought that Spinosaurus had the ability to swallow large fish and other prey items, with the ability to prey upon pterosaurs and juvenile dinosaurs.[11]
Isotope analysis and taphonomic evidence both support the use of aquatic resources by Spinosaurus.[12][13]
Carved reconstructed of the cranium of Spinosaurus, held by paleoartist Tyler Keillor.
Competition:
Interestingly, large crocodylomorph remains (e.g. Sarcosuchus) dominate Aptian beds in Africa and South America, but are unknown from the Cenomanian of North Africa. The largest crocodylomorph from the latter beds is Elosuchus; with a skull ~100 cm long, it would have been smaller than the giant spinosaurids. Larger crocodylomorphs seem rare in the Kem Kem Beds and Bahariya fauna. This suggests that spinosaurids became freshwater apex predators in North Africa as giant crocodylomorphs declined. Spinosaurids may have either replaced a vacant niche left behind by these crocodylomorphs or actively outcompeted them into extinction.[14]
References:
[1] paleobiodb.org/classic/basicTaxonInfo?taxon_no=53372
[2] Evers SW, Rauhut OWM, Milner AC, McFeeters B, Allain R. 2015. A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco. PeerJ 3:e1323 doi.org/10.7717/peerj.1323
[3] GIMSA, J., SLEIGH, R., & GIMSA, U. (2016). The riddle of Spinosaurus aegyptiacus’ dorsal sail. Geological Magazine, 153(3), 544-547. doi:10.1017/S0016756815000801
[4] Hone, D., & Holtz, T. (2021). Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?. Palaeontologia Electronica.
[5] Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D. M., Zouhri, S., Myhrvold, N., & Iurino, D. A. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science (New York, N.Y.), 345(6204), 1613–1616. doi.org/10.1126/science.1258750
[6] Ibrahim, N., Maganuco, S., Dal Sasso, C. et al. Tail-propelled aquatic locomotion in a theropod dinosaur. Nature 581, 67–70 (2020). doi.org/10.1038/s41586-020-2190-3
[7] Larramendi, A., Paul, G. S., & Hsu, S. Y. (2020). A review and reappraisal of the specific gravities of present and past multicellular organisms, with an emphasis on tetrapods. Anatomical record (Hoboken, N.J. : 2007), 10.1002/ar.24574. Advance online publication. doi.org/10.1002/ar.24574
[8] Foffa, D., Cuff, A. R., Sassoon, J., Rayfield, E. J., Mavrogordato, M. N., & Benton, M. J. (2014). Functional anatomy and feeding biomechanics of a giant Upper Jurassic pliosaur (Reptilia: Sauropterygia) from Weymouth Bay, Dorset, UK. Journal of anatomy, 225(2), 209–219. doi.org/10.1111/joa.12200
[9] Lemanis, R., Jones, A. S., Butler, R. J., Anderson, P. S., & Rayfield, E. J. (2019). Comparative biomechanical analysis demonstrates functional convergence between slender-snouted crocodilians and phytosaurs (No. e27476v1). PeerJ Preprints.
[10] Cuff, A. R., & Rayfield, E. J. (2013). Feeding mechanics in spinosaurid theropods and extant crocodilians. PloS one, 8(5), e65295. doi.org/10.1371/journal.pone.0065295
[11] Hendrickx, C., Mateus, O., & Buffetaut, E. (2016). [url=https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144695Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa[/url]. PloS one, 11(1), e0144695. doi.org/10.1371/journal.pone.0144695
[12] Amiot, R., Buffetaut, E., Lécuyer, C., Wang, X., Boudad, L., Ding, Z., ... & Zhou, Z. (2010). Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods. Geology, 38(2), 139-142.[13] Beevor, T., Quigley, A., Smith, R. E., Smyth, R. S. H
[13] Ibrahim, N., Zouhri, S., & Martill, D. M. (2021). Taphonomic evidence supports an aquatic lifestyle for Spinosaurus. Cretaceous Research, 117, [104627]. doi.org/10.1016/j.cretres.2020.104627
[14] Arden, T. M., Klein, C. G., Zouhri, S., & Longrich, N. R. (2019). Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurids. Cretaceous Research, 93, 275-284.